Introduction
It is well-known that cancer driver genes play an
important role in the occurrence and development of tumors. Gene
mutations continuously accumulate with aging or in tumor lesions.
However, previous studies have found that there are also numerous
somatic mutations in normal adult tissues, as well as specific
‘cancer-driven mutations’, for example, mutations in the
cancer-driving gene Notch1 (1,2).
Therefore, there is a view that the main risk factor for cancer
development is not persistent carcinogenic mutations, but the
exposure of the initial mutant cells to an environment that
promotes tumor development (3,4). This is
consistent with the decisive role of the tumor microenvironment in
the process of tumor formation and development. Data analysis found
that the telomeric repeat-binding factor (TRF)2 protein was
dysfunctional, possessing the dual characteristics of promoting
cell mutation and the formation of extracellular tumor-promoting
environment factors, and plays a key role in the process of tumor
formation (5–8).
Telomeres are a type of DNA-protein complex
structure, which seal the ends of the chromosome arms of eukaryotic
cells and maintain the stability of the genome. Telomeres are
essential for the integrity of the chromosomes. In all mammals,
telomeres are composed of a highly conserved hexamer (TTAGGG)
tandem repeating DNA sequence and telomere binding protein, which
is composed of the shelterin complex, accessory factors and
telomerase (5,9,10). The
shelterin complex can specifically bind to telomere DNA to cause
the telomere end structure to form a telomere loop (T-loop),
assisting the telomeres to form caps and protecting the ends of
chromosomes from DNA double-strand break (DSB) recognition, thereby
inhibiting DNA damage response (DDR), maintaining the integrity of
telomere structure and function, and preserving chromosome
stability (9). Human shelterin
complexes consist of six core proteins: TRF1 and TRF2,
TERF1-interacting nuclear factor 2 (TINF2), TRF2-interacting
protein 1 (also known as repressor/activator protein 1; RAP1),
protection of telomeres 1 (POT1) and POT1-binding protein 1 (also
known as telomere-binding protein POT1-interacting protein 1; TPP1)
and a tripeptide kinase (10). The
shelterin proteins, TRF1 and TRF2, display specific affinity for
the double-stranded (ds)DNA of the telomere, while the POT1-TPP1
shelterin subcomplex covers single-stranded telomere G-rich
overhangs (11). Based on the
currently available-related research, the present review aimed to
systematically discuss the basic structure and functional
characteristics of TRF2, and elucidate its role in tumor
formation.
TRF2 structure and function maintenance
TRF2 is an important polypeptide in the telomere
shelterin complex. The gene is located on chromosome 16q22.1, 30 kb
in length and is widely expressed in numerous tissues in humans
(National Centre Biotechnology Information via flatfile form;
ftp://ftp.ncbi. nih.gov/genomes/). TRF2 includes 4 functional
domains, which are described as follows: i) The N-terminal basic
region [glycine and arginine-rich (GAR) or basic domain], which is
rich in glycine and arginine residues, and is indispensable for the
basic protective function of TRF2 (9). Knocking down expression of non-coding
telomere repeats-containing RNA (TERRA) using short inhibiting RNA
lead to telomere dysfunction, including increased metaphase
chromosomal aberrations (an 1.4-fold increase in telomere repeat
signal intensity, but no significant change in average telomere
length) and telomere lesions associated with p53-binding protein 1
(53BP1) and phosphorylated histone H2AX (γ-H2AX) (9,12). The
TRF2 basic domain can recruit and bind TERRA independent of the
telomere sequence, and it can interact with subunits of the origin
recognition complex 1 domain associated with the TRF2 basic domain
to promote telomere heterochromatin formation and participate in
the regulation of DDR (12). The
complete basic domain, via non-specific electrostatic attraction,
notably enhances the affinity of TRF2 to DNA and protects the
telomere displacement loop (D-loop) from cleavage by endonucleases
(13). ii) The dimerization domain,
also known as the TRF homology (TRFH) domain, is involved in the
formation of homodimers. Similar to TRF1, it has the same α helix
structure, resembling a twisted horseshoe (5). The dimerization interface exhibits
unique interactions, which can prevent heterodimerization (5). TRFH is a protein domain that can
recognize specific peptides of the (Y/F)XL motif, and it is also a
hub structure of multiple (Y/F)XL motif-containing protein signals,
which mediate telomere length control and end protection. For
example, the TRF2-TINF2 interaction regulates telomere formation
and activation of telomerase (a specialized DNA polymerase which
adds repeats of telomere DNA to the ends of chromosomes) (14,15). In
addition, the TRF2-Apollo and TRF2-microcephalin interactions
regulate DNA damage repair response (16,17) and
play an important role in molecular dimerization, DNA binding and
telomere positioning (18). In the
TRFH domain of TRF2, lysine and arginine residues are surrounded by
90 bp DNA. This structure is important in maintaining the
topological structure of telomere DNA, inhibiting the activation of
the ataxia telangiectasia-mutated (ATM) kinase checkpoint and
preventing telomere non-homologous end joining (NHEJ) (19). iii) The orientation sequence (nuclear
localization signal), or flexible hinge domain, contains TRF2 and
other shelterin proteins, such as RAP1 and TINF2 interaction sites
(11,20). iv) The C-terminal domain
[Myb/homeodomain-like telobox DNA-binding domain (DBD)] is mainly
responsible for the specific recognition and binding of the telobox
DNA repeat sequence, TTAGGG (14,21). TRF2
DBD binds to telomere DNA and nuclear chromosome arrays, and rarely
non-specifically binds to non-telomere DNA sequences (22).
Telomeres functionally maintain the stability of
chromosomes and the biological clock-like function of DNA
replication, and their stability depends on 2 mechanisms. The first
relies on telomerase or alternative lengthening of telomere (ALT)
activity, which can compensate for replication erosion; the second
relies on its own special chromatin organization, which protects
the ends of chromosomes from abnormal signals and repair (9,11).
Chromatin-mediated telomere protection involves several
protein-regulated pathways, which directly or indirectly bind to
telomere DNA, such as the shelterin complex (23,24). The
TRF2 protein in the shelterin complex is important for maintaining
the normal functional state of telomeres and is considered as a
telomere protective protein. For example, TRF2 folds human
telomeres into loops to prevent unnecessary DDR, blocks ATM
signaling and NHEJ, and promotes telomere replication, serving as a
key regulator of telomere integrity (9,24–26). The structure and main functions of
TRF2 are shown in Fig. 1.
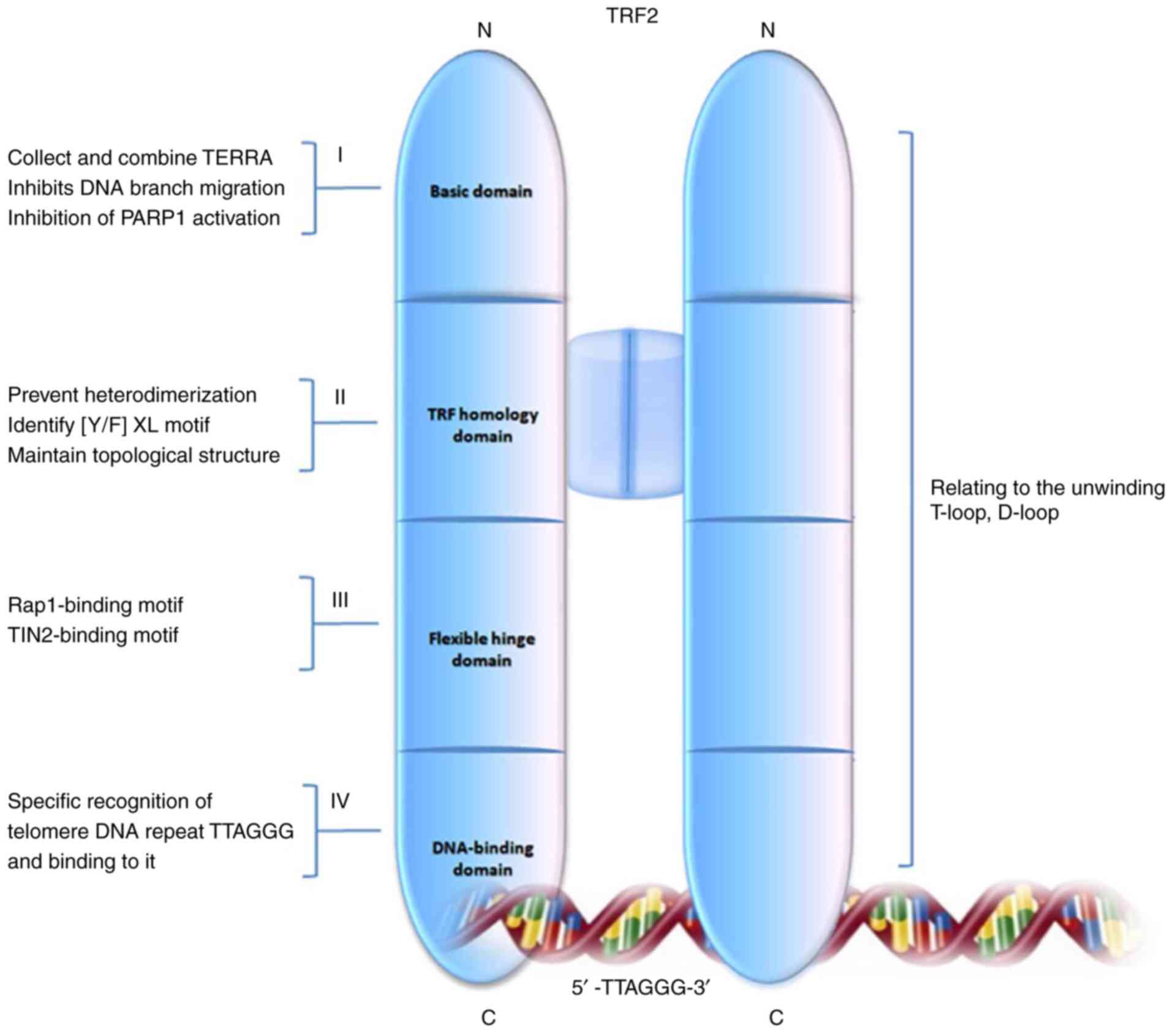 | Figure 1.Schematic diagram of TRF2 structure
and its main functions. TRF2 includes 4 functional domains: Basic,
TRF homology, flexible hinge and DNA-binding domains. Different
structural domains have functional characteristics of I, II, III
and IV, respectively. The various structural domains of TRF2 act
synergistically and play an important role in regulating the
orderly assembly and untwisting of the T- and D-loops. The
structural variation of TRF2 can cause large fragments of telomeres
to be lost, resulting in severe telomere shortening. TRF2,
telomeric repeat-binding factor 2; T-loop, telomere loop; D-loop,
telomere displacement loop; C, C-terminal; N, N-terminal; TERRA,
telomeric repeat-containing RNA; RAP1, repressor/activator protein
1; TINF2, TERF1-interacting nuclear factor 2. |
Functional characteristics of TRF2 and the
maintenance of cell chromosome stability
TRF2 maintains and dynamically regulates the
structure of eukaryotic telomere end loops. TRF2 protein mutation
or abnormal expression (particularly domain mutation associated
with loop assembly, including the basic and the RBM domains) leads
to loss of the T-loop, loss of protection of the DNA end, DDR or
end-to-end fusion, chromosomal instability (CIN), cell aging,
apoptosis or malignant transformation. With respect to morphology,
the linear chromosomes of eukaryotes end with nuclear protein
complex telomeres. Mammalian telomere DNA is composed of several
kbp 5′-TTAGGG-3′ repeats and the ends are terminated in a 50–400
nucleotide single-stranded overhang, with a G-rich sequence. The
dsDNA of telomere repeats forms a T-loop and the 3′ end of the
single strand of the telomere is inserted into the dsDNA of the
telomere (i.e., the 3′ overhang end is paired with the C-rich
strand). The G-rich strand, in the double strand, is replaced to
form a replacement loop, which is called D-loop. This telomere ring
structure hides the ends of the chromosomes to protect the
stability of telomere DNA from DDR or end-to-end fusion (27).
With respect to biological function, TRF2 mediates
the formation of loop structures. The functional elimination of
TRF2 in the cell will result in the loss of the loop structure
(28). In the absence of other
shelterin proteins, TRF2 can also form a T-loop alone (29), mediating telomere protection at
different cell cycle stages (30).
The mechanism of T-loop formation is associated with the ability of
TRFH domains in TRF2 to bind DNA with low affinity and change the
DNA topology (19). It has been found
that the TRF2 dimer, via several lysine and arginine residues
located around TRFH, encapsulates ~90 bp DNA and forms DNA
topology. For TRF2 mutants with insufficient encapsulates (named
Top-less), the topological structure of telomere DNA can be changed
and the number of terminal T-loops can be reduced (19). This encapsulation also only occurs
when the Myb/SANT domain of TRF2 and the dsDNA TTAGGG repeat
sequence are bound to each other (19).
The formation and/or maintenance of the T-loop does
not require the N-terminal basic domain of TRF2, but this domain
can bind to and cover the branched-DNA 3-way junction. The 3-way
junction structure contains the poly(ADP-ribose) polymerase (PARP)1
activation site, and the activation of PARP1 can cause the harmful
poly(ADP-ribosyl)ation (PARsylation) of the telomere protein
(31). The TRF2 protein basic domain,
H31A and the TINF2 protein jointly inhibit PARP1 activity, and
inhibit the migration of branched-DNA to form double Holliday
junction (dHJ), stabilize the T-loop, prevent it from being cut and
thereby prevent telomere shortening (31). Therefore, the basic domain of TRF2 is
crucial for the stability of the T-loop.
In the cell cycle, the T-loop is a highly dynamic
and regulated structure, and its assembly and disassembly are
coordinated in the cell cycle. The T-loop requires twining and
unfolding in an orderly and alternate manner, controlling the
protection and replication functions of the DNA ends, and
coordinating the structural contradiction between the two functions
(32). A previous study found that
there was a cyclin-dependent kinase (CDK)2 phosphorylation site at
Ser365 of TRF2, which was shown to be phosphorylated in the
G1, G2 and M phases, but not in the S phase
(32). Due to the dephosphorylation
of the phosphatase, PP6R3, which is enriched in the S phase, a
narrow window is created at Ser365. The helicase regulator of
telomere elongation helicase 1 (RTEL1) can enter and untie the
T-loop instantaneously, relieving the collision between the S-phase
replication fork and the T-loop, and promote telomere DNA
replication. At the Ser365 site of TRF2 in cell cycle phases, other
than the S phase, RTEL1 is repelled and released from telomeres,
and TRF2 is again phosphorylated, restoring the T-loop structure.
This not only prevents the disorderly unwrapping of the T-loop and
inappropriate activation of ATM, but also counteracts the
replication conflicts of the secondary DNA structure in the
telomere and the genome. Therefore, the phosphorylation switch in
TRF2 coordinates the assembly and disassembly of the T-loop during
the cell cycle, protecting telomeres from replication stress and
unplanned DDRs (32).
In-depth studies have found that the interaction
site of RTEL1 with TRF2 is mapped to an uncharacterized C4C4
metal-binding motif (33). The
p.R1264H mutation in the C4C4 motif can disrupt the S-phase
RTEL1-TRF2 interaction and leads to a T-loop split, accompanied by
rapid changes in telomere length, telomere loss and T-circle (TC)
formation. The amino acid p.I124D mutation, in the TRFH
dimerization domain of TRF2, can also specifically disrupt the
binding to RTEL1 and prevent its recruitment on telomeres, thereby
phenotypically replicating the p.R1264H mutation in RTEL1, leading
to T-loop abnormal resection, uneven telomere length, telomere loss
and TC formation (33). Therefore,
TRF2 plays an important role in coordinating T-loop assembly and
disassembly during the cell cycle.
In addition to the T-loop, the D-loop must also be
disassembled immediately during DNA replication. An in vitro
purified protein function test found that TRF2 exerted a notable
inhibitory effect on single-stranded (ss) DNA strand invasion and
D-loop formation mediated by Rad51 in the telomere template, which
is not conducive to the formation of the D-loop structure; however,
in the non-telomeric region of the genome or Myb domain mutants, it
could promote single strand invasion and D-loop formation, and was
conducive to homology recombination, which may be associated with
the specific structure and function of the dsDNA-binding domain
(Myb SANT) of TRF2 (34).
After the formation of the D-loop, TRF2 binds to
DNA, the toughness of its basic domain decreases, and it becomes
structural and rigid. It can also prevent telomere D-loop unwinding
caused by a variety of DNA unwinding factors and ensure the
stability of the T-loop (13). RAP1
is recruited to the telomere and binds to the RBM domain of TRF2 to
form the RAP1-TRF2 complex, which can restore the toughness of the
TRF2 basic domain, eliminate non-specific DNA and histone
electrostatic affinity of the TRF2 N-terminal, inhibit its
interaction with DNA and mediate chromatin loosening (13). At this time, despite the presence of
TRF2, RAP1 can also promote D-loop unwinding with helicase, which
is beneficial for telomere extension and chromosome end
replication. It can be seen that the combination of RAP1 and TRF2
can inhibit the affinity of the TRF2 basic domain to telomere DNA
and eliminate the D-loop structure (13). Based on the aforementioned mechanism,
as the cell cycle changes, TRF2 dynamically adjusts the D-loop to
alternately unwind and close the loop, which not only stabilizes
telomeres, but also ensures the normal progression of the cell
cycle.
In summary, the different modes of action and
functional state of the telomere protein, with TRF2 as the core,
determines the loading and unloading regulation of the T- and
D-loops to maintain chromosome stability, and the cell cycle of
normal cells.
TRF2 inhibits telomere DDR and abnormal
repair
TRF2 regulation of DDR
Telomere deprotection occurs in aging or damaged
cells, which coincides with the natural erosion of the chromosome
ends. When the telomere reaches a very short length, DDR is no
longer under control. The metabolic activity continues; however,
the cell will still enter an irreversible stagnation state
(35). The mechanism is associated
with the decrease of TRF2 protein content and function. When the
telomere is severely shortened, it will erode numerous TRF2-binding
sites. Insufficient TRF2 recruitment will lead to the failure of
telomere protection, loss of its inhibitory effect on ATM and ATM
activation. Phosphorylated ATM can induce subsequent localization
of DDR components. For example, γ-H2AX and 53BP1 are recruited to
the ends of the chromosomes, leading to the formation of what is
referred to as telomere dysfunction-induced foci (TIF), activating
the DDR cascade and inducing telomere repair (36).
The inhibitory effect of TRF2 on ATM has been
associated with the functions of its TRFH domain and hinge domain,
in the following 2 forms: i) The TRFH domain of TRF2 promotes the
formation of the T-loop structure, hiding the 3′ ssDNA at the end
of the telomere, in the double-stranded structure, which does not
produce an interaction between the MRN complex and the DNA end
similar to the DSB sensor; thus, preventing MRN-dependent ATM
activation (35–37). Since the maintenance of the T-loop
structure is associated with the functional integrity of TRF2, TRF2
dysfunction can induce the loss of T-loop protection at the
telomere ends and expose the DNA, which can trigger the initial
steps in the DDR pathway, trigger the ATM cascade and produce DDR
(19). ii) The inhibitory DDR
response (IDDR) domain of TRF2 regulates the activity of downstream
proteins in the ATM cascade. At the γ-H2AX-labeled telomere, TRF2
relies on the deubiquitinating enzyme, BRCC3, and the ubiquitin
ligase, UBR5, via the ‘IDDR motif’ in its hinge domain to hinder
the recruitment of E3 ubiquitin ligase, RNF168, and inhibit its
activity (37). With respect to
RNF168, the DNA damage signal cascade is interrupted to prevent the
positioning of 53BP1, thereby preventing chromosome end-to-end
fusion (37). The ability of TRF2 to
prevent the activation of the DDR pathway can also be enhanced with
other shelterin subunits. For example, TRF1 recruits TINF2, which
is conducive to the effective combination of TRF2 and telomeres,
and strengthens the telomere protection function of TRF2. TINF2,
together with TRF1 and TRF2, improve the telomere positioning and
function of TRF2 (38,39). A previous study found that Top-less
telomere TIF cells maintained a certain NHEJ inhibitory ability,
indicating that there was another mechanism protecting the ends of
telomeres from NHEJ (19). If the
expression of the TRF2-interacting protein, RAP1, in the telomeres
is eliminated at this time, the repressive ability of NHEJ will be
reduced and end-to-end fusion will occur. It can be observed that,
when topology-mediated telomere protection is compromised, the
presence of RAP1 can also serve as a backup mechanism for
preventing NHEJ (19).
TRF2 has an inhibitory effect on the DDR cascade. It
is well-known that the DDR cascade, under the monitoring of cell
cycle checkpoints, can block the cell cycle, facilitate the repair
of damaged DNA and maintain chromosome stability (30). Cells that are difficult to repair due
to severe damage are eliminated by initiating the apoptotic
cascade, to avoid the proliferation of abnormal cells with unstable
chromosomes due to telomere damage, which will cause tissue and
organ dysfunction (30,31). TRF2 can prevent abnormal DNA enzymatic
excision and maintain the integrity of telomeres and chromosomes
(31). However, under certain
conditions, TRF2 also exerts a significant inhibitory effect on
DDR, that damages or severely shortens telomeres, allowing cells to
evade cycle checkpoint monitoring and repair. Chromosomally
unstable cells grow and proliferate under the promoting effect of
cyclin. Due to CIN, the mutant cells in the genome cannot be
cleared in a timely manner and mutations accumulate, increasing the
risk for the occurrence of tumors and other genetic diseases (such
as xeroderma pigmentosum syndrome) (32,33,35,40).
The inhibitory effect of TRF2 on the DDR cascade following telomere
injury is shown in Fig. 2.
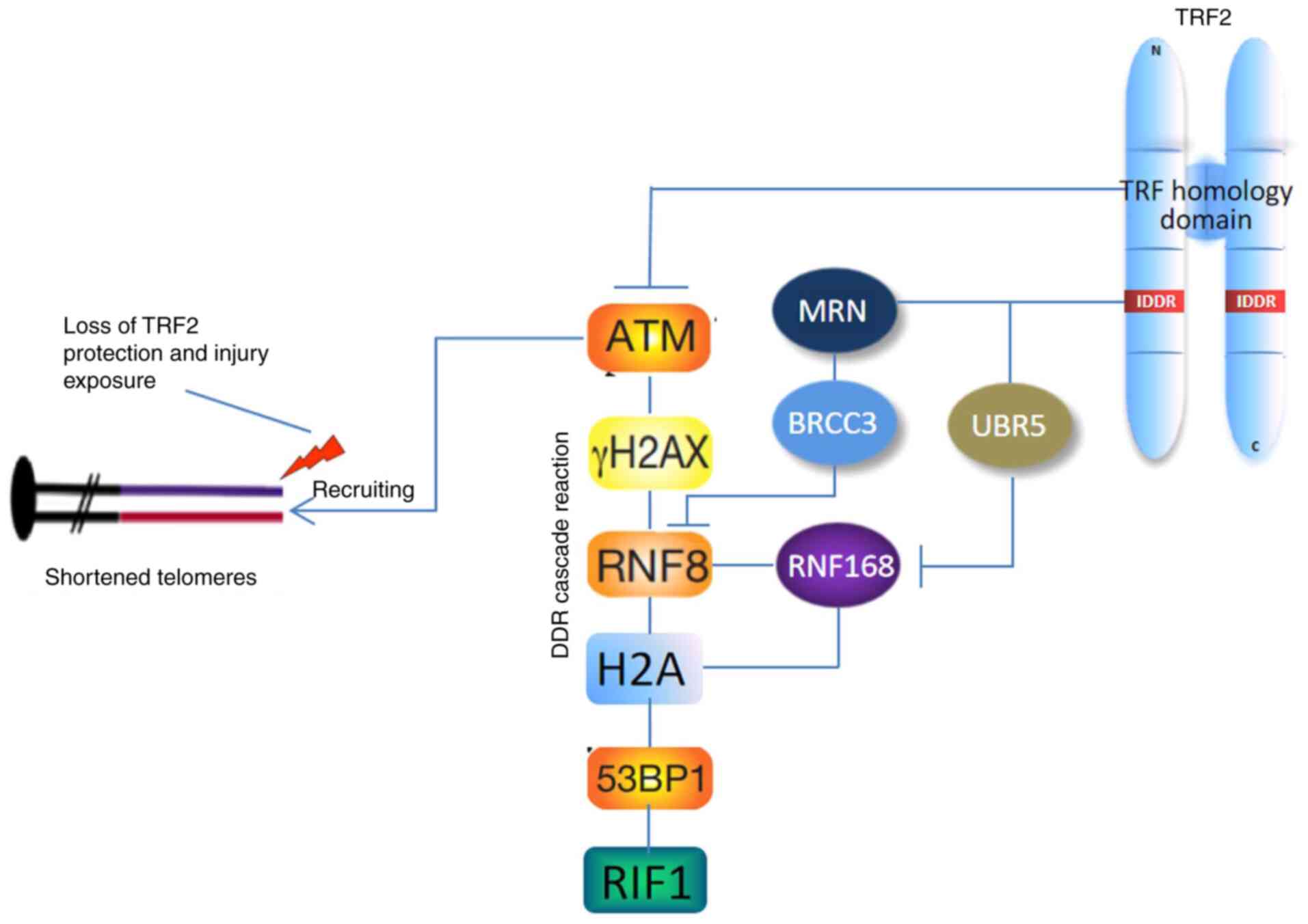 | Figure 2.Schematic diagram of the inhibitory
effect of TRF2 on the DDR cascade following telomere injury. The
TRFH domain of TRF2 can directly inhibit ATM. The IDDR domain of
the flexible hinge region can inhibit and degrade RNF8 and RNF168
via BRCC3 and UBR5, respectively, and arrest the DDR cascade at the
RNF168 level. TRF2, telomeric repeat-binding factor 2; TRFH, TRF
homology; DDR, DNA damage response; IDDR, inhibitory DDR response;
ATM, ataxia telangiectasia mutated; γ-H2AX, phosphorylated H2AX;
H2A, H2A histone; 53BP1, p53 binding protein 1; RIF1, replication
timing regulatory factor 1; C, C-terminal; N, N-terminal. |
Regulatory effect of TRF2 on the abnormal repair of
telomere ends. When cell chromosomes are severely shortened, due to
replicative wear and tear, or other pathological factors (such as,
abnormal expression of TRF2, XPF enzyme digestion or telomere
replication excision), the end loses the protection of the
shelterin complex protein and is recognized by the cell as broken
DNA (36,37). At this time, DDR is activated, which
induces abnormal repair and the mutual combination of broken ends,
forming multiple centromeric chromosomes, losing the ability of
normal replication and division of cells, and finally being
eliminated (9,29,33,35). This
process may also be considered as the physiological effect of the
self-protecting mechanism of the body, by eliminating aging or
mutated cells, to maintain the relative chromosome stability of
living tissue and cells, and avoid the occurrence of tumors or
other related diseases, such as Bloom's syndrome or Werner syndrome
(41,42). Similar to the DSB repair mechanism,
telomere ends are usually repaired in three ways: Canonical NHEJ
(C-NHEJ), alternative NHEJ (A-NHEJ) and homologous recombination
repair (HDR). Among these, C-NHEJ is the most common repair method.
After TRF2expression is suppressed, the telomere ends lose their
protective structure. Under the induction of DDR, the circular
nucleoprotein Ku70/80 heterodimer is loaded to the telomere ends to
prevent free DNA from being decomposed by nuclease (43,44). After
the protein, Ku recruits DNA-dependent protein kinase catalytic
subunits, it further recruits and phosphorylates several factors,
such as DNA ligase (Lig) IV and XRCC4, and finally each protein
contributes to the completion of telomere-telomere fusion (30,43,44). It
was previously demonstrated that the Ku70 N-terminal Ku70α5 region
residues contributed to the Ku-Ku association between telomeres to
form heterotetramers and prepared DNA end-joining. TRF2 can also
interact with Ku via the Ku70α5 domain (the functional area of TRF2
has not yet been determined), blocking Ku from connecting the 2
telomere ends to directly inhibit NHEJ at the telomeres (45). Therefore, TRF2 is considered as the
main inhibitor of telomere C-NHEJ, which directly inhibits C-NHEJ
via the negative regulation of Ku (45). The inhibition of C-NHEJ by TRF2 is
shown in Fig. 3. Fig. 3A shows that the damaged shortened
telomere ends bind to the Ku70/80 protein, and via the mutual
affinity of the Ku70 5α domains, a Ku70/80 heterotetramer is
formed, leading to chromosome end-to-end fusion. Fig. 3B shows that TRF2 directly binds to the
5α domain of Ku70 to prevent the formation of Ku70/80
heterotetramers and inhibit the recruitment to the telomere ends of
DNA-PKcs, Lig4, XRCC4 and XLF making the ends unable to be
effectively connected.
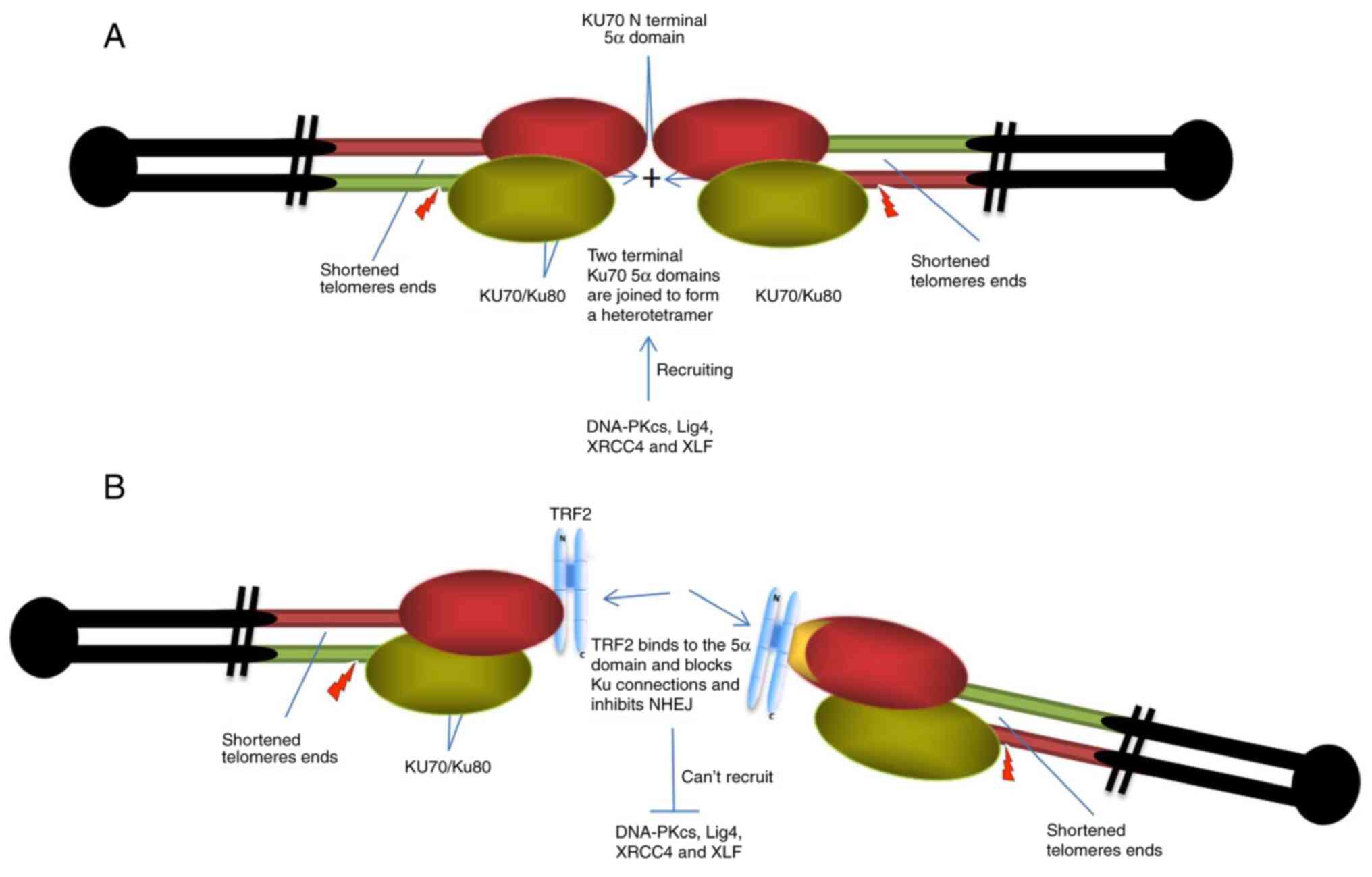 | Figure 3.Inhibitory effect of TRF2 on damaged
telomere C-NHEJ. (A) The damaged shortened telomere ends bind to
the Ku70/80 protein, and via mutual affinity of the Ku70 5α
domains, a Ku70/80 heterotetramer is formed, leading to chromosome
end-to-end fusion. (B) TRF2 directly binds to the 5α domain of Ku70
to prevent the formation of the Ku70/80 heterotetramers and
inhibits the recruitment to the telomere ends of DNA-PKcs, Lig4,
XRCC4, XLF, making the ends unable to be effectively connected.
TRF2, telomeric repeat-binding factor 2; C-NHEJ, canonical
non-homologous end joining; DNA-PKcs, DNA-dependent protein kinase
catalytic subunits; Lig4, DNA ligase IV; XLF, XRCC4-like
factor. |
The normal telomere T-loop structure, mediated by
TRF2, can also limit the loading of the Ku70/80 heterodimer at the
telomere ends; therefore, the presence of TRF2 at the telomeres may
be beneficial for eliminating C-NHEJ. As the telomere C-NHEJ fusion
requires ATM kinase activation (24),
the key component of the cascade is 53BP1. It was previously found
that, in the absence of 53BP1, even without TRF2, telomere C-NHEJ
is very rare (46). On the one hand,
the accumulation of 53BP1 at the telomeres, by recruiting a series
of 53BP1-dependent DDR factors, including RIF1 and related shieldin
complexes, reduces the end DNA strand excision effect of multiple
nucleases, and inhibits other methods of telomere end joining, such
as DNA homologous recombination repair (47). On the other hand, 53BP1 can promote
the dynamic movement of DNA damage sites in the nucleus.
Unprotected telomeres and other sites of DNA damage become more
mobile and move in a larger area of the nucleus. 53BP1-mediated
mobility increases the chance of telomeres approaching another
telomere, thereby facilitating telomere fusion (47). Furthermore, the presence of 53BP1 can
also cause the aggregation of unprotected telomeres, which may also
promote telomere fusion (48). As
aforementioned, the regulation of 53BP1 by the IDDR domain of the
TRF2 hinge region can effectively inhibit C-NHEJ-mediated terminal
fusion; therefore, inhibition of C-NHEJ by TRF2 is more conducive
to aging or abnormal cells bypassing the checkpoint block,
promoting cell cycle progression and causing pathological mitosis,
adversely affecting chromosome stability (49).
When cell telomeres lack Ku or Lig4 nucleoproteins,
another alternative DSB repair mechanism is often used, referred to
as A-NHEJ (50,51). At the end of the telomere, PARP1 binds
to the 5′ ds-ss transition site and is activated, thereby
initiating A-NHEJ. The PARsylation of PARP1 to nearby proteins
(including histones) creates a binding platform for DNA Lig3 and
DNA polymerase. If the ends of 2 different DNAs have 3′ overhangs
and exhibit slight homology, DNA polymerase-mediated filling and
Lig3 (or replication ligase Lig1) end ligation will result in
A-NHEJ. The homology required for A-NHEJ is minimal (≥1 base pairs)
(50,51). In the case of 2 dysfunctional
telomeres, the homology between the 3′ overhang ends (2 bp per
telomere repeat) is sufficient for A-NHEJ. Ku70/80 in C-NHEJ
usually inhibits the A-NsHEJ pathway by competing with PARP1 at the
end (50,51). When shelterin is completely removed
from the telomeres of cells lacking Ku70/80, almost all telomeres
are fused by A-NHEJ (50). Another
study found that, as TRF2 is an independent factor in the formation
of the T-loop, it may hide the telomere ends in the T-loop to
inhibit PARP1 activation, and A-NHEJ does not occur. However,
knocking down the expression level of TRF2 with short inhibiting or
shRNA from Ku70/80-deficient cells will only result in a mild
telomere fusion phenotype (51).
Mouse embryonic fibroblasts lacking TPP1 or POT1a/b can also
exhibit low levels of A-NHEJ fusion (52), and TINF2 can also limit the
accumulation of PARP1 in telomeres and inhibit the initiation of
A-NHEJ (31), indicating that other
shelterin components can synergistically enhance the inhibition of
A-NHEJ. Therefore, mechanistically, T-loop formation and PARP1
blocking may be the 2 main mechanisms in which telomeres avoid this
pathway. In addition, it was found that the Werner syndrome (WRN)
protein exerts a regulatory effect on the induction of C-NHEJ and
A-NHEJ. WRN promotes C-NHEJ via its helicase and nuclease
functions, and prevents 5′ end excision in a non-catalytic manner
by inhibiting MRE11 and CTIP, thereby inhibiting A-NHEJ (53).
The physiological significance of cell replicative
senescence is the inhibition of tumor formation. In critical short
telomeres lacking telomerase, the HDR between sister chromatids can
partially compensate for the loss of telomere repeats when
telomerase is missing, thereby promoting early onset of aging
(54), which can also be considered
as the 3rd method of telomere terminal repair. In human cells,
telomere HDR activation is usually inhibited by Ku70/80, but in the
absence of some subunits of shelterin, such as POT1, very low
levels of telomere sister chromatid exchange can be detected even
in the presence of Ku70/80 (55). The
key to HDR activation is the activation of PARP1, and a variety of
mechanisms that inhibit PARP1 activity can prevent HDR. Rai et
al (56) found that the shelterin
substructure, RAP1, and the basic domain of TRF2 can act together
to inhibit the localization of PARP1 to telomeres, thereby
inhibiting the telomere recruitment of the HJ dissociation enzyme,
SLX4, and ultimately inhibit HDR. In the absence of the basic
domain of TRF2 and the RAP1 protein, PARP1, SLX4, and other
proteins involved in HDR (for example, Mus81 and Sgs1), promote the
rapid resection of T-loop structure telomeres, telomere loss, and
the formation of chromosome ends without telomeres, eventually
leading to an extensive telomere-free chromosome fusion in virus
specific T cells (56). With respect
to the related mechanism, as the local loss of TRF2 activates
PARP1, the PARsylation of PARP1 can lead to the recruitment of the
SLX4-SLX1-Mus81 HJ dissociation enzymes (57). These HJ dissociation enzymes may
cleave the T-loop 3-way connection, but will not cause loss of
telomeres or untie the T-loop, which promotes HDR. However, in the
absence of the TRF2 basic domain, the 3-way connection can freely
branch and migrate, thereby potentially forming a dHJ (31). Treatment of dHJ by HJ dissociation
enzymes may result in the excision of telomere DNA in the loop. The
TRF2 basic domain can bind to the 3- or 4-way connection of DNA.
This binding is not related to the DNA sequence and has similar
affinity for these 2 structures (dissociation constant, ~200 nM)
(31). TRF2 binds to the 3-way
connection at the base of the telomere T-loop, which can mask the
PARP1 activation site at the 5′ concave end, inhibits the
recruitment of SLX4 and other endonucleases (Mus81), and also
stabilizes the 3-way connection, inhibits branch migration, avoids
the formation of dHJ, and ultimately prevents T-loop cleavage
(31). However, investigation of this
mechanism did not reveal that RAP1 has a synergistic effect with
TRF2 in inhibiting PARP1 or T-loop cleavage (54), which is inconsistent with the
aforementioned results, and further research is required (31).
In summary, following telomere damage and
shortening, the DDR cascade is activated, and the 3 end-repair
methods, C-NHEJ, A-NHEJ and HDR may be triggered, according to the
characteristics of the internal environment of the cell nucleus.
This abnormal repair leads to telomere dysfunction and CIN.
Surviving cells change with the cell cycle, and abnormal
replication and mitosis will produce a variety of random DNA
recombination mutations.
Regulation of telomere length by
TRF2
Telomere length is associated with chromosome
stability. Telomeres are severely shortened and lose the protective
effect of shelterin substructures, which can cause CIN, and
abnormal expression of the telomere protein can also cause changes
in telomere length, which also affects chromosome stability
(58). TRF2 is a key player in
telomere protection and telomere length maintenance. It can prevent
loss of T-loops and is also a negative regulator of telomere
length. For example, the overexpression of TRF2 leads to shortening
of telomeres in normal primary fibroblasts and telomerase-positive
lung, liver and gastric cancer cells (59–61).
Previous studies have found that the mechanism of TRF2 and cell
telomere length regulation is associated with the following
aspects. i) On the one hand, as aforementioned, the interaction of
the N-terminal basic domain of TRF2 with the dissociation
enzyme-related protein, SLX4, and different types of endonucleases
involved in dissociation activities, such as GEN1 and MUS81,
prevent the cutting of telomeres (62). Furthermore, various types of
dissociation enzyme, such as SLX4 and GEN1 activities are checked
via this domain to promote the precise repair of stalled forks,
which occur during telomere replication (62). On the other hand, the TRF2
homodimerization domain can recruit SLX4, which is also a
structure-specific DNA repair nuclease scaffold, and can recruit
the Holliday ligation enzyme, SLX1 and other nucleases (GEN1 and
MUS81) to the telomeres to prevent telomere damage, lengthening and
fragility, and plays multiple roles in regulating telomere
homeostasis (63). ii) TRF2 promotes
telomere shortening via the DNA repair endonuclease XPF-excision
repair cross complementation group 1 (ERCC1) enzyme complex.
XPF-ERCC1 serves as a structure-specific endonuclease, involved in
nucleotide excision repair, cross-linking repair and DNA
recombination (61,64,65). In
the absence of the XPF-ERCC1 nuclease, K5-TRF2 mice, which
overexpress TRF2, did not exhibit significant telomere shortening,
suggesting that the TRF2-bound XPF enzyme rapidly degrades
telomeres in the presence of increased TRF2 expression (61). Therefore, telomere shortening promoted
by TRF2 requires the function of XPF-ERCC1. An in-depth study has
demonstrated that the nuclease activity of XPF-ERCC1 was essential
for nucleotide excision repair, but the XPF-ERCC1 mutant protein,
without nuclease activity, could also induce TRF2-mediated telomere
shortening, revealing that the role of XPF-ERCC1 was unrelated to
its nuclease function, and this phenomenon was conserved between
mice and humans; therefore, the specific mechanism of action
requires further investigation (64).
XPF-ERCC1 can also negatively regulate the binding of TRF2 and
telomere DNA. Introducing wild-type XPF into XPF-deficient cells
(human Phoenix andGM08437B cells) was shown to reduce the
association between TRF2 and telomere DNA by >40%. If the
function of its nuclease domain is still lost, the aforementioned
effect can be eliminated. These findings indicated that XPF-ERCC1
controls TRF2 and telomere length maintenance via 2 unique
mechanisms (65). iii) The
TRF2-mediated growth factor heregulin (HRG)β2 is a negative
regulator of telomere length. Co-immunoprecipitation and imaging
experiments demonstrated that HRGβ2 could be partially localized at
the end of chromosomes, bind to and interact with RAP1/TRF2
(66,67). Overexpression of HRGβ2 promoted
notable upregulation of TRF2 protein expression, while knockout of
the protein markedly reduced the protein expression level of TRF2
(66,67). In non-invasive, HRG-negative MCF-7
breast cancer cells, engineered overexpression of the HRGβ2 subtype
resulted in notable shortening of the telomeres; in highly
invasive, HRG-overexpressing MDA-MB-231 and Hs578T cells,
antisense-mediated HRGβ2 inhibition could increase telomere length
(66,67). Therefore, the telomere length
regulation function of HRGβ2 may be associated with its regulation
of TRF2 (66,67). iv) TRF2 can regulate the expression of
human telomerase reverse transcriptase (hTERT), a telomerase
rate-limiting catalytic protein component. Telomerase is an
important factor in the regulation of telomere length, but it is
mostly in a state of expression inhibition in somatic cells
(68–70). A previous study found that subtelomere
genes were regulated by the telomere length-dependent loop
(telomere position effect over long distances; TPE-OLD), which
silences subtelomere genes by affecting telomere position (68). The TERT gene is a megabase in
length, from the end of the human 5p chromosome. Cells with long
telomeres at the end of the 5p chromosome form a telomere chromatin
loop in the hTERT site domain, which inhibits hTERT
expression (68). It was found that
TRF2 knockdown notably reduced the interaction between telomere and
the hTERT locus, suggesting that TRF2 may be involved in the
T-loop interaction on the hTERT locus (68). With respect to the mechanism, with
cell aging or telomere shortening and decreased TRF2 level, the
chromatin ring and hTERT locus could become separated, and
notable epigenetic changes, such as DNA methylation and histone
modification, may occur in the hTERT promoter region, which
may activate hTERT transcription and enhance telomerase
activity (68). A previous study
further demonstrated that there were frequent interstitial
telomeric DNA sequences (ITS) downstream of the TERT site in
higher primates (69). Through the
interaction with interstitial TTAGGG repeats, TRF2 may be
responsible for the interaction between telomere and hTERT
ITS in cells with long telomeres, and act on the hTERT
promoter to inhibit its expression. When TRF2 protein expression
level decreases, the interaction between T-loop and ITS is
weakened, which may relieve the inhibition of hTERT, induce
the expression of hTERT and affect the telomerase activity
assembly (69).
TRF2 extratelomeric epigenetic
regulation function
TRF2 can bind to a specific sequence of dsDNA via
its C-terminal Myb domain. In addition, there is another broader
non-ds binding method. Studies have found that, in the human
genome, TRF2 has thousands of binding sites outside the telomeres
and is associated with the whole genome (70,71). This
association enables TRF2 to participate in extratelomeric DNA
repair and gene transcription regulation (71–73). The
repetitive guanine-rich sequence can form a 4-stranded DNA tetramer
structure in solution (74–77) and TRF2 is one of the few proteins with
specific high affinity for this structure (78,79). This
affinity requires the N-terminal basic domain of TRF2 and the
C-terminal DNA-binding domain to collaborate, and its affinity is
stronger than the specific sequence of dsDNA (73). Therefore, a large part of non-telomere
TRF2-binding sites are composed of non-ds G-quadruplexes (G4s). The
G4 motif is a DNA secondary structure. The core structure of the G4
motif comprises stacked planar tetrads of guanine residues
stabilized by Hoogsteen base pairing (74). It is considered to be a global gene
regulatory motif and this function requires interaction with a
transcriptional regulatory protein specifically associated with
G4s. G4s are abundant in gene promoters and are evolutionarily
conserved in a variety of animals and microorganisms, such as
human, chimpanzee, mouse and rat, and several kinds of bacteria
(Escherichia coli and Saccharomyces cerevisiae)
(74,75,80). There
are extensive TRF2-G4 associations in the entire genome and they
regulate gene expression in a G4s motif-dependent manner (70). The interaction between non-telomere
TRF2 and promoter G4s induces gene regulation via epigenetic
modification, leading to the activation or silencing of histone
marker genes. It has been demonstrated that, following binding of
TRF2, histones are modified, resulting in transcriptional
inhibition of 8 out of 9 genes, changing the expression and
epigenetic status of the target promoter (70).
p21 (also known as CDKN1A), a CDK inhibitor, is
negatively regulated by TRF2 in a G4-dependent manner, i.e., loss
of TRF2 can enhance the activation of p21 (72). This is achieved by TRF2 by recruiting
the REST-co-REST-LSD1 inhibitory complex. The activation of p21
leads to cell cycle arrest and apoptosis/senescence, which is the
key mechanism underlying the action of DNA damage-inducing
anticancer drugs. In cancer cells (HT1080 fibrosaroma and
MDA-MB-231 breast cancer cell lines) with relatively higher TRF2
protein expression levels, p21 activation was found to be notably
reduced following drug treatment, suggesting that it is associated
with drug resistance (72).
The G4 structure, −600 bp upstream of the
hTERT promoter, determines the occupancy rate of TRF2 in
this promoter. Following TRF2 knockdown, the TRF2 occupancy rate of
G4s in the promoter decreases, which can induce the transcriptional
activation of hTERT (81).
This is due to the polycomb inhibitor polycomb repressive complex 2
(PRC2) in the hTERT promoter being dependent on TRF2
recruitment. PRC2-induced H3K27 inhibits histone-tagged
trimethylation, leading to the inhibition of hTERT. The G to
A mutation in the cancer-specific and highly recurrent hTERT
promoter disrupts the TRF2-G4 interaction. This mutation has been
associated with several types of cancer, including glioblastoma,
melanoma, hepatocellular carcinoma and urothelial carcinoma
(81–85). It destroys the G4 structure in the
hTERT promoter, inhibits the binding of TRF2 to the promoter
and eliminates the inhibitory effect of the PRC2, resulting in the
reactivation of hTERT. Therefore, the epigenetic regulation
of hTERT, caused by the interaction of TRF2-G4, may be of
notable value for the treatment of various types of cancer
(81). In addition, there are
numerous epigenetic modifications in tumor regulatory genes that
cancer cells rely on for survival, all of which are associated with
the TRF2 occupancy rate of the G4 structure in their respective
promoters, such as c-MYC, KRAS and VEGF (86–89). This
shows that, apart from telomeres, TRF2 is crucial for the
expression and epigenetic modification of tumor regulatory genes. A
previous study found that the epigenetic status and expression of
numerous TRF2 target promoters, located at a great distance from
telomeres, are sensitive to telomere length (90). The TRF2 occupancy rate of long
telomeres at remote genomic sites is usually low. On the contrary,
when the telomeres are relatively short, the non-telomere TRF2
binding increases notably (90). This
telomere length-sensitive promoter TRF2 occupancy rate changes,
resulting in epigenetic modification of the promoter. A previous
study referred to this as a telomere segregation and partition
(TSP) model, which can affect telomere-dependent gene expression in
the whole genome (91). TSP is
different from the TPE-OLD model, as the TPE-OLD model only affects
genes ~10 Mb from the telomere (91).
With telomere shortening, the distribution of TRF2 inside and
outside the telomere changes correspondingly. Under the combined
action of related cofactors or gene mutations, senescent cells
exhibit dual characteristics of activation or inhibition of
transcription epigenetic modification, which eventually leads to
the inhibition of tumor suppressor genes or the promotion of
oncogene expression.
In summary, in mammalian cells, TRF2 is notably
associated with telomere length, involving a variety of regulatory
mechanisms and markedly affecting gene expression. Chromosome
stability depends on the telomere structure of sufficient length.
Within a certain range, the abundance of telomere binding protein
is positively correlated with the length of telomere DNA repeats.
An increase or decrease of TRF2 expression level can cause changes
in telomere length. When the critical length of telomeres is
reached, causing CIN, the genome is prone to mutations affected by
a variety of environmental factors for example, ionizing radiation
and mutagenic compounds. When affected by telomerase or selective
telomere lengthening mechanisms, telomeres can maintain their
critical length and prevent further shortening. This hypothesis is
consistent with the fact that tumor cells have short telomeres. At
this time, under the telomere protection or extra-telomeric
regulation of TRF2, pathological responses, such as DDR or
end-to-end connections, that induce cell proliferation arrest or
apoptosis, cannot be evoked, which maintains the cell cycle,
promotes cell proliferation and growth, and generates immortal
cells. As these cells grow and proliferate, gene mutations
gradually accumulate, which may eventually induce the development
of a variety of tumors.
Association of TRF2 dysfunction with CIN and
tumor formation
CIN refers to the increase in the frequency of
chromosome gain or loss in the process of cancer cell division
compared with that in normal cells, which leads to the decrease in
the stability of the genome and induces a variety of mutations,
such as polyploid or aneuploid formation. CIN is one of the
hallmarks of cancer and is attributed to errors in chromosome
separation during mitosis (92,93). CIN
plays an important role in cancer by accelerating the accumulation
of genetic changes responsible for the transformation of cancer
cells and is the main driving factor behind tumor development
(92–94). CIN may occur through a variety of
mechanisms, including cell cycle regulation, DDR, DNA replication,
chromosome segregation and telomere dysfunction.
Telomeres form protective caps at the ends of linear
chromosomes to prevent nuclear degradation, end-to-end fusion,
irregular recombination and CIN, and protect them from loss of
genetic information and inappropriate handling when DNA is damaged.
Telomeres are crucial for preserving the integrity and stability of
chromosomes (49,95). It is hypothesized that tumor formation
is suppressed by 2 replicative senescence and crisis (96). Under normal genetic and epigenetic
background conditions, telomeres shorten with each cell division
(physiological telomere wear), and finally stop cell proliferation
by activating DDR, which regulates the life span of somatic cells
and limits their renewal ability (96). If cells lose the checkpoint function,
the ability to protect the ends of linear chromosomes from
end-to-end fusion is eventually lost. A previous study revealed
that the state of the telomere complex and the length of the
telomere sequence was associated with chromosome stability
(97). Specifically, telomere complex
protein damage or abrasion telomere shortening activated THE ATM
and ATR kinases via unprotected chromosomal ends, induced the DDR
cascade, and caused cell cycle arrest. During this period, cells
try to repair the damage. If the DNA damage cannot be repaired, the
senescence response is triggered, which exerts an antitumor effect
(98,99); on the other hand, the loss of telomere
protection and cycle checkpoint blockade may lead to a telomere
crisis, which is characterized by further shortening of
dysfunctional telomeres and extensive chromosomal fusion. Cells in
this critical state will produce extensive apoptosis (100). End-to-end fusion of chromosomes
during the telomere crisis can cause spontaneous mitotic arrest.
During the period of spontaneous mitotic arrest during the telomere
crisis, telomere deprotection is a potential molecular signal
leading to cell death. The destruction of TRF2 leads to rapid
mitotic cycle stagnation or cycle extension, which is an important
signal leading to cell death (101).
Since most cell deaths before the crisis are associated with
mitosis arrest, it is hypothesized that prolonging mitosis is the
main mechanism that restricts senescence and prolongs cell lifespan
(101). Telomere crisis represents a
highly unstable state of the genome and entering the
break-fusion-bridge (BFB) cycle via end-to-end chromosome fusion is
a classic mechanism underlying telomere-driven genome instability.
The dysfunctional telomeres fuse with the ends of other chromosomes
to produce dicentric chromosomes to initiate the anaphase
bridge-dependent cycle (6,102,103).
Anaphase chromatin bridges may or may not be broken, leading to
numerous structural rearrangements, segment duplication or abnormal
separation of polyploid chromosomes (103). Chromosomal rearrangements commonly
associated with telomere dysfunction include irreversible
translocation, region amplification and segment deletion, and
non-segregating events that lead to aneuploidy. These dicentric
chromatids formed by the replication and fusion of sister
chromatids will participate in the fusion bridge breaking cycle
until new telomeres are added to the free ends, leaving multiple
segments of DNA amplification and deletion accumulation (103). Segment duplication may lead to
increased expression of genes located in the amplified region,
which is one of the mechanisms underlying oncogene activation. This
can also confer resistance to chemotherapeutic drugs by increasing
the number of copies of genes encoding drug targets (103). In brief, these rearrangements can
produce chimeric genes or oncogene disorders, as well as changes in
gene dosage required to induce tumorigenesis. In the absence of
functional tumor suppressor genes, they can promote cancer
occurrence (104–106). Therefore, CIN promotes the malignant
transformation of precancerous cells at the genetic level. With the
delayed activation of telomerase or alternative elongation
mechanisms, precancerous cells can achieve unlimited replication
capacity and immortality and, ultimately, complete cancerous
transformation (Fig. 4).
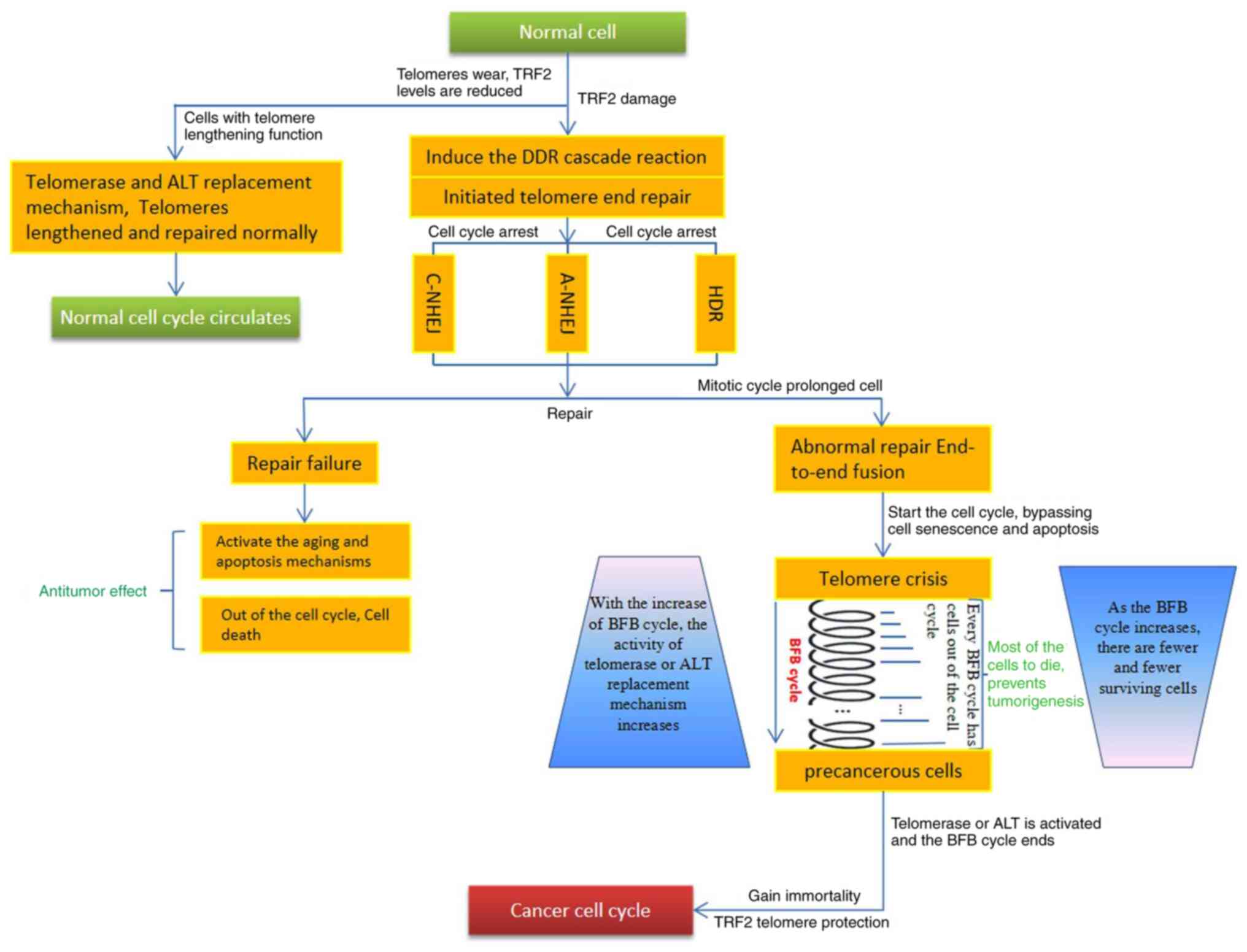 | Figure 4.Abnormal function of telomere TRF2
leads to the transformation of normal cells into tumor cells. The
chromosomal telomeres of normal cells display shortening, TRF2
localization and functional damage due to replicative senescence
and other causes such as Drugs, radiation damage, and induce DDR
and abnormal telomere end repair. A proportion of the cells that
fail to repair are eliminated due to the initiation of the
senescence and apoptosis mechanisms. Another proportion of the
cells, due to the prolonged mitotic cycle, bypass senescence and
apoptosis, and are abnormally repaired, exhibiting telomere end
fusion and entering a telomere crisis period. During this crisis
period, the chromosomes are extremely unstable and are reorganized
on a large scale, which manifests as the BFB cycle. During each BFB
cycle, most of the cells die, which can reduce the number of
precancerous cells. A small number of surviving cells exhibit
gradually activated telomerase and ALT telomere substitution
activity due to a variety of factors (such as genetic mutations),
resulting in telomere extension and ending of the BFB cycle. TRF2
binds the repeat sequence on the lengthened telomere, protects the
newly formed telomere and reduces chromosomal instability. At this
time, the recombinant cells (precancerous cells) become immortal
and evolve into cancer cells. TRF2, telomeric repeat-binding factor
2; DDR, DNA damage response; BFB, break-fusion-bridge; ALT,
alternative lengthening of telomere; C, canonical; NHEJ,
non-homologous end joining, A, alternative; HDR, homologous
recombination repair. |
TRF2 plays a crucial role in regulating molecular
events, which maintain telomere integrity (107). The abnormally high or low expression
level of TRF2 may lead to CIN and induce fatal tumor changes. In
some types of cancer (such as breast cancer, stomach cancer and
hematological malignancies), TRF2 expression is reduced (108–110).
With respect to the underlying mechanism, TRF2 knockdown could
destroy the T-loop structure and deprive telomere ends of
protection. SLX4, GEN1 and MUS81 lack TRF2 regulation, which can
cause telomere shortening (62); in
addition, it has been demonstrated that the amino-terminal basic
domain of TRF2 binds to the histone H2A isotype protein, H2ac, and
recruits H2ac to the telomeres to participate in the regulation of
telomere structure and/or function (111). The loss of H2ac may lead to the
accumulation of telomere-related ATM-dependent DNA damage factors
in H2ac-deficient cells, DDR, rapid loss of telomere repeats and
G-rich strand loss (111). Telomere
loss or damage can induce dysfunction, trigger DNA damage signal
transduction, block cell cycle progression and induce cell aging
and apoptosis. Some mutant cells can bypass cycle arrest, enter
telomere crisis, accelerate improper repair, produce chromosome
end-to-end fusion, initiate the BFB cycle, cause genomic
instability and promote tumorigenesis (101). It was found that TRF2 knockdown
could induce the expansion and metastasis of cancer stem cells
(CSCs), while reducing the number of CD34+ stem cells (112,113).
This reveals that the change in telomere length regulation mediated
by TRF2 deficiency has a profound impact on stem cells (40,114).
This effect involves epithelial-to-mesenchymal transition (EMT)
caused by the expansion of CSCs (115), as well as the overexpression of
telomerase and the ALT mechanism, which promotes the formation of
metastatic tumors (116,117). The genetic and biological
characteristics of these TRF2 deletion-mediated metastatic CSCs
require further research for characterization.
The overexpression of TRF2 may also play an
important role in the fate of cancer cells. Notable increases in
TRF2 protein expression levels have been reported in a group of
human cancers (such as epithelial carcinogenesis, gastric cancer
and liver cancer) (118–120). The overexpression of TRF2 could
accelerate the telomere erosion rate of human primary cells (UMUC3
bladder cancer cells) in the absence of telomerase, and even
trigger DDR (121,122). A previous study (123) found that most chromosomal fusions
induced by TRF2 overexpression were accompanied by extensive
deletions involving subtelomere regions of chromosomes, causing
sustained DNA replication stagnation, and the chromosomal ends were
severely shortened randomly, whereas the fusion of these
chromosomes usually contained microhomologous regions of 1 to 6
nucleotides. The mechanism may be as follows: TRF2 overexpression
leads to the formation of excessive compact DNA-protein complexes
and the binding of the TRF1 protein, which promotes telomere
replication, to telomeres decreases, thereby hindering the
development of telomere replication forks, leading to telomere DNA
replication stagnation. The resulting decomposition of telomeric
ultrafine anaphase bridges results in the random loss of large
fragments of telomere sequences and subtelomere regions. Then, via
the selective A-NHEJ repair process, the broken chromosomes are
fused, which eventually leads to CIN (123,124).
In addition, as aforementioned, the telomere shortening promoted by
TRF2 also requires XPF-ERCC1 cooperation, which is unrelated to its
endonuclease function, and the mechanism underlying its association
with TRF2 overexpression leading to replication fork arrest
requires further in-depth investigation. Another study demonstrated
that TRF2 was overexpressed in colorectal cancer tissues and in the
SW480 cell line, and the mechanism was associated with the
transcription factor, Sp1, which is involved in the upregulation of
TRF2 expression (125). It has also
been demonstrated that the promoter of TRF2 is rich in GC sequences
(70-75%), and there was no notable TATA box. Sp1 is a multi-gene
family member with sequence-specific DNA binding to the C-terminal
zinc finger motif (126–128). It activates transcription by binding
to the GC/GT box and plays a key role in the regulation of gene
expression in mammals lacking the TATA box (126,127).
Sp1 regulates cancer-related targets, including PDEGF, p21, VEGF,
TGFβ, cyclin D1, E2F1, c-Fos, TGF-α and osteopontin, which are key
transcription factors caused by cancer (128–130).
Therefore, it was hypothesized that, in certain types of tumor
cells (such as human colon adenocarcinoma WiDr and Caco-2 cell
lines), Sp1 may also upregulate the expression of TRF2 and induce
tumor development (129,130). In addition to Sp1, β-catenin has
also been shown to activate TRF2 transcription. The overexpression
of the transcription factor complex β-catenin/T-cell factor (TCF)1E
leads to an increase in TRF2 protein expression level and its
binding to telomeres to confer telomere protection (131). By evaluating the number of TIF,
i.e., the number of telomere foci co-localized with 53BP1 immune
signal, it was confirmed that changes in β-catenin expression
affected telomere protection (131).
In addition, it was recently demonstrated that the expression level
of TRF2 and TRF1 could also be regulated by the chromatin
remodeling complex, BRM-SWItch/sucrose non-fermentable (SWI/SNF)
(132). BRM could be recruited to
the promoters of TRF2 and TRF1, which provides a basis for
effective transcription, and also raises the possibility that
several other identified or unknown factors may collaborate with
BRM-SWI/SNF to regulate the transcription of TRF2 and TRF1, whereas
its deletion reduces the mRNA and protein expression levels of TRF2
and TRF1 (132). In addition, the
TERF2 gene has a typical CpG island around its transcription
start site. The hypomethylation of the TRF2 gene promoter and the
CpG island of exon 1 is one of the important mechanisms underlying
TRF2 gene upregulation, and hypomethylation may be involved in the
upregulation of TRF2 in gastric cancer (133). The microRNAs (miR) miR-23a and
miR-155 inhibited the translation of TRF2 and TRF1 by targeting the
3 untranslated regions of their transcripts (134,135).
A schematic diagram of the factors involved in the
TRF2 tumor formation-promoting effect is shown in Fig. 5.
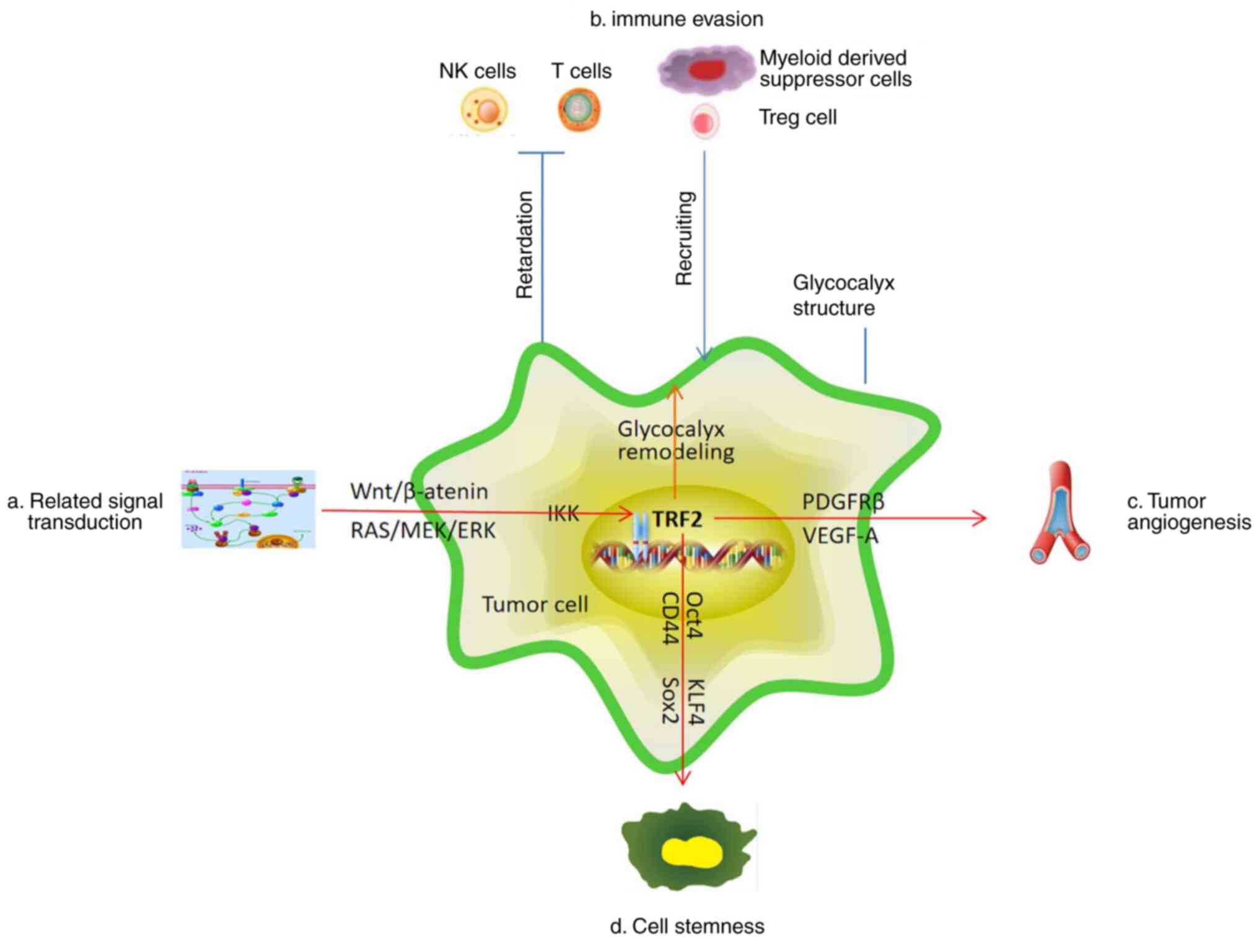 | Figure 5.Schematic diagram of the TRF2-tumor
promoting effect. (a) Transduction via signaling pathways, such as
Wnt, RAS and IKK, upregulates, activates and stabilizes TRF2
molecules in tumor cells, and stimulates internal and external
regulatory mechanisms that are conducive to tumor development via
regulation of epigenetic changes or gene expression. (b) TRF2
reconstitutes the glycocalyx via gene regulation of glycocalyx
components outside the cell membrane, thereby recruiting
myeloid-derived suppressor cells, suppressing immune effector
cells, and producing immune escape. (c) TRF2 promotes the formation
of vascular endothelial cells in the tumor cell microenvironment
and promotes tumor angiogenesis via the regulation of tumor
angiogenesis factor expression. (d) TRF2 maintains the
characteristics of tumor stem cells by regulating the gene
expression and stability of tumor cell stemness-related
transcription factors and related proteins, thereby promoting tumor
cell proliferation and cancer metastasis. TRF2, telomeric
repeat-binding factor 2; IKK, IκB kinase; NK, natural killer; Treg,
regulatory T cells. |
TRF2 and tumor angiogenesis
The role of TRF2 in cancer depends not only on its
protective effect on telomeres, but also on its ability to regulate
gene expression. Angiogenesis is important for the progression of
malignant tumors. A previous study found that TRF2 and Wilms' tumor
suppressor 1 (WT1) were highly expressed in the blood vessels of
tumors in patients with prostate, pancreas, breast, lung and kidney
cancer, and TRF2 co-localized with WT1 in vascular endothelial
cells (7); however, it was not
expressed in the blood vessels of healthy adjacent tissues, further
proving that TRF2 was the transcription target of WT1. WT1 binds to
and activates the TRF2 promoter in human endothelial cells to
promote TRF2 expression. TRF2 can stimulate the migration,
proliferation and formation of tube-like structures by endothelial
cells, thereby increasing tumor angiogenesis (7). Knockdown of TRF2 could suppress the
angiogenic properties of endothelial cells. The specific mechanism
could be that TRF2 binds to and transactivates the gene promoter of
platelet-derived growth factor receptor β (PDGFRβ) in tumor
endothelial cells, which promotes PDGFRβ expression on the tumor
endothelial cell membrane, thereby enhancing the formation of the
tumor vascular network and promoting tumor growth (7). This effect was independent of the
telomere-protective effect of TRF2 (7). Another study found that TRF2 could
notably affect the expression level of VEGF-A in the ‘secretory
body’ of cancer cells (136). There
was a positive correlation between high levels of TRF2 and VEGF-A,
which promoted endothelial cell differentiation and angiogenesis,
and it may be used as a new type of tumor prognostic marker to
identify high-risk colorectal cancer subgroups (136). The underlying mechanism could be due
to the combination of TRF2 and distal regulatory elements, which
can promote the expression level of sulfatase 2 (SULF2), an
endoglucosamine-6-sulfatase, and is a heparan sulfate proteoglycan
(HSPG) catabolic enzyme. By inducing the post-synthesis
modification of HSPGs, TRF2 weakens HSPGs ability to bind and
isolate signaling molecules containing heparin-binding domains,
including VEGF-A, thereby inhibiting the binding of VEGF-A to the
plasma membrane, which can affect tumor vascularization and,
consequently, tumor growth and metastasis (137). Clinically, TRF2/SULF2 expression
level may be a biomarker of poor prognosis for patients with
colorectal cancer (CRC) (137).
Therefore, compared with the classic antitumor neovascularization
therapy, using TRF2 as a therapeutic target may represent a major
development in CRC therapy.
TRF2 and tumor signal transduction
pathways
TRF2 expression is regulated by the
Wnt/β-catenin signaling pathway
As aforementioned, the Wnt/β-catenin signaling
pathway activates the expression of TRF2 and maintains the level of
TRF2 that is essential for telomere protection in human cancer
cells, normal cells and mouse intestinal tissues (131). The canonical Wnt signaling pathway
activates and stabilizes β-catenin, translocates to the nucleus,
then combines with members of the TCF-lymphoid enhancer factor
(LEF) transcription factor family to activate gene transcription
(131). Analysis of the human
TERF2 gene using the Ensembl genome database (http://grch37.ensembl.org/index.html)
revealed a potential regulatory region (termed Reg) downstream of
the transcription start site (131).
The TERF2-Reg sequence contains 6 putative TCF-LEF transcription
factor-binding sites. The overexpression of β-catenin alone, or the
co-overexpression of β-catenin and TCF-LEF induced specific
activation of the natural Reg region (131,138).
The expression level of β-catenin also determines the expression
level of the RAP1 gene encoding the TRF2-interacting protein. These
results indicated that β-catenin regulated telomere protection by
regulating the level of TRF2. Using gene knockout mice, reducing
the expression of the β-catenin gene, Ctnnb1, could trigger
an increase in telomere TIF, which was accompanied by a decrease in
cell viability and an increase in the percentage of cells
expressing β-galactosidase (a marker of cellular senescence)
(131). Typically, in normal and
cancer cells, either in mice or humans, the activation of
Wnt/β-catenin signaling leads to an increase in TRF2 expression
levels and enhanced telomere protection (131,138),
which is also one of the important mechanisms of abnormal
proliferation of tumor cells.
TRF2 stability in cells is regulated
by the RAS/RAF/MEK/ERK signaling pathway
In addition to playing a key role in cancer cells,
the RAS/RAF/MEK/ERK signaling pathway also represents one of the
most important pathways required for growth control, normal
development, differentiation and environmental response in the
majority of tissues. Throughout the cell cycle, subtle regulation
of ERK1/2 activity is important for G1/S and
G2/M transition (139,140).
ERK1/2 activity decreases in the S phase compared with that in the
G1 and M phases; however, a basic ERK1/2 activity
continues throughout the cell cycle (141). TRF2 expression levels and telomere
protection are regulated by MAP kinase signaling. Compared with
that in the precursor cell line, the expression level of the TRF2
protein is increased in immortalized cells (142). In a variety of human cancers (such
as epithelial carcinogenesis, gastric cancer and liver cancer), the
expression of TRF2 at the RNA and protein level increases (8,121), and
its internal mechanism may be associated with the regulation of
TRF2 amino acid residue phosphorylation. As the serine 323 (Ser323)
residue of TRF2 is embedded in the MAPK consensus PXSP
phosphorylation motif, ERK1/2 and TRF2 interact in different types
of cells (including normal as well as cancer cells) (143,144).
TRF2 Ser323 can be phosphorylated by ERK1/2, which increases the
half-life and stability of TRF2, participates in telomere
protection, and promotes tumor growth by maintaining TRF2 levels
(143,144). The mutant non-phosphorylated form of
TRF2 is unstable and alters telomere termination in a dominant
negative manner, triggering telomere uncovering, growth arrest and
tumor reversion. Therefore, the constitutive activation of MAPK
signal transduction in cancer leads to an increase in TRF2 levels
and telomere protection (143,144).
These results suggested that telomere stability is dependent on
TRF2 phosphorylation, which is regulated by the Ras/Raf/MEK/ERK
axis (144).
TRF2 is affected by IκB kinase (IKK)
regulation in the signaling pathway
IKKα is known to be a key regulator of
tumorigenesis by affecting the NF-ĸB signaling pathway (145). The activation of IKK-β has been
associated with cancer pathogenesis and inflammatory diseases
(146). Research on liver CSC
revealed that, when the inflammation-related genes, IKKα and IKKβ,
were both overexpressed, the interaction between the telomere DNA
probe and TRF2 notably increased (146,147).
In addition, the loading of POT1, Exo1 and SNM1b on the telomere
DNA also increased, and the telomere length notably increased. On
the contrary, when a single IKKγ was overexpressed, the interaction
between the telomere DNA probe and TRF2 was markedly reduced, the
loading of POT1, Exo1 and SNM1b on the telomere DNA was also
reduced, and the telomere length was notably reduced (146,147).
Mechanistically, IKKα, IKKβ and IKKγ depended on HP1 to change the
methylation of histone H3 on lysine 27 (H3K27me3). IKKα and IKKβ
enhanced, whereas IKKγ inhibited, the competition among HP1α, HP1β
and HP1γ. Therefore, IKKα and IKKβ inhibit, whereas IKKγ enhances,
the activities of H3K27 methyltransferases, SUZ12 and EZH2,
regulating the methylation of H3K27 in the promoter region of
HOTAIR. H3K27me3 negatively regulated the expression level of
HOTAIR. HOTAIR exerted carcinogenic effects of IKKα, IKKβ and IKKγ
in liver CSCs, and promoted the increase in TRF2 and the
accumulation of telomere length (146,147).
Collectively, these findings indicated that IKKα and IKKβ
increased, whereas IKKγ decreased, telomere length and the
expression levels of TRF2 and other related protective proteins
[POT1, phosphorylated (p)POT1, Exo1, pExo1, SNM1B,
pSNM1B/CST-AAF]in liver CSCs, which jointly regulated the
proliferation ability of CSCs (147).
In summary, TRF2 exhibits a variety of network
connections with tumor regulatory signaling pathways, allowing
tumor cells to achieve the networked regulation of multiple
signaling pathways of TRF2 protein. Telomere function status, gene
expression and functional proteins in the cytoplasm coordinate to
promote rapid tumor cell proliferation, invasion and migration,
immune evasion and drug resistance.
TRF2 and CSCs
CSCs are important for the continuous growth,
recurrence and metastasis of malignant tumors. Telomere protection
protein is an important maintenance component of the biological
characteristics of CSCs and the role of TRF2 is particularly
important. Short telomeres lacking TRF2 protection can hinder the
proliferation of stem cells, affect their tissue regeneration
potential and trigger the development of age-related diseases
(105). The existence of CSCs
mediated by TRF2 has been associated with the drug resistance,
invasion and metastasis of cancer syndrome treatment (105).
Tumor radiotherapy can cause the formation of DSB
in irradiated cells and trigger DDR. When DSB cannot be repaired
effectively, the cells clear themselves through programmed
apoptosis (148). CSCs usually
exhibit high TRF2 expression level, which maintains the globular
phenotype of tumor stem cells cloned in vitro (149,150).
It was previously demonstrated that, after radiation treatment, the
expression level of TRF2 and RAP1 in the stem cell cloning sphere
increased, and the interaction between the 2 could also regulate
the activity of hTERT, which was essential for the
maintenance of telomeres (151).
Enhanced expression of TRF2, RAP1 and hTERT in stem cell
clone spheres could improve telomere DNA repair and protection, and
counteract genomic instability caused by telomere shortening or
DSB, thereby effectively resisting cell death induced by DNA damage
caused by radiotherapy, chemotherapy and other factors (such as
aging), maintain the survival of tumor cells, and produce tolerance
to treatment (152).
A feature of the ALT mechanism in tumor cells is
the assembly of ALT-associated promyelocytic leukemia-nuclear
bodies (APBs) on the telomeres. The formation of APBs can induce
telomere aggregation, telomere compaction and the concomitant
depletion of the shelterin protein TRF2, strongly enrich the
phosphorylation of ATM kinase and induce DDR (152,153).
Therefore, APBs induce ATM phosphorylation by changing the state of
telomere chromatin; thus, inducing DDR and recombinant repair of
ALT-positive tumor cells and promoting telomere maintenance
(153). TRF2 can regulate cell
proliferation and differentiation in APBs by interacting with
repressor element 1-silencing transcription factor (REST) via a
non-telomere effect. The TRF2-REST complex may play a role in
protecting REST from ubiquitin proteasome degradation, thereby
maintaining CSCs in an undifferentiated stage (154). Glioblastoma contains self-renewing
glioblastoma stem cells (GSCs), which are usually resistant to
chemotherapy and radiotherapy. GSCs highly express REST, which may
promote their resistance to standard cancer treatments (155,156).
TRF2 stabilizes telomeres and REST, and maintains the self-renewal
of neural stem cells and GSC (157).
Viral vector-mediated infection using short hairpin (sh)RNAs
targeting TRF2 mRNA could reduce TRF2 and REST expression levels in
the GSCs of patients with glioblastoma, resulting in a decrease in
GSC proliferation, while the protein expression levels of L1CAM and
t3-tubulin, which are normally expressed by neurons after mitosis,
increases, suggesting that the loss of TRF2 leads to the
differentiation of GSCs (157). The
loss of TRF2 also renders GSCs sensitive to temozolomide, a DNA
alkylating agent used in the treatment of glioblastoma (157). Using TRF2 as a target was shown to
notably improve the survival rate of GSCs in a xenograft mouse
model (157). These findings
indicate a role of TRF2 in GSCs, which includes maintaining
stability, ensuring proliferation and chemotherapy resistance,
suggesting that TRF2 could be a potential therapeutic target for
glioblastoma (157).
Knocking down the expression level of TRF2 could
also inhibit the migration potential of oral cancer cell lines, and
at the same time reduce the expression of key factors that maintain
CSCs in the Cal27 cell line and the generated oral squamous cell
carcinoma cells, including CD44, Oct4, Sox2, KLF4, c-Myc, β-catenin
and hTERT molecules (150). Further
research found that TRF2 has a strong affinity and interaction with
the CSC marker, KLF4, which may be an important mechanism for the
maintenance of CSCs, indicating that TRF2 was associated with the
transmission of CSC-like phenotypes in oral cancer cells (150).
From the aforementioned results, it may be inferred
that TRF2 plays a key role in maintaining the function of CSCs.
TRF2 elimination in CSCs could further promote differentiation,
loss of the biological characteristics of stemness, increased
sensitivity to drugs or radiotherapy, inhibition of EMT, invasion
and metastasis of tumor cells, which may improve the efficacy of
clinical tumor treatment and long-term prognosis.
TRF2 and tumor immunity
In human malignant lesions, the increase of TRF2
expression level in some tumor cells (such as epithelial
carcinogenesis, gastric cancer and liver cancer) could reduce the
recruitment and activation of natural killer (NK) cells, and assist
the tumor cells to bypass innate immune surveillance. This is a key
step in human tumorigenesis, and it shows that TRF2 controls the
biological functions of NK cells and affects tumorigenesis in a
DDR-independent manner (8,158). Mechanistically, high TRF2 expression
level was found to be associated with the expression level of 3
genes (HS3ST4, GPC6 and VCAN) involved in the
biosynthesis of HSPG. These genes all contain an ITS as a binding
site for TRF2 (158,159). After binding, TRF2 induces changes
in the chromatin structure at the ITS, becomes a TRF2-dependent
enhancer element, and activates the expression of neighboring genes
(159). The promoting effect of TRF2
on the expression level of the HSPG gene was crucial to the
formation of the polysaccharide calyx outside the cytoplasmic
membrane, acting as a general remodeling agent of the glycocalyx,
as the overexpression of TRF2 changed its overall structure,
increased its stiffness and shortened its length (159). Changes in glycocalyx structure
change the micro-gradient of cytokines, chemokines and growth
factors involved in the recruitment and function of immune cells,
which is required for myeloid-derived suppressor cell (MDSC) tumor
recruitment. The proteoglycan version encoded by VCAN acts
as a toll-like receptor (TLR)2 ligand, and the TLR2 signal
activates the immunosuppressive function of MDSCs via the IL-6
autocrine pathway, i.e., IL-6/JAK1/2/STAT3 signaling induces the
immunosuppressive autocrine circuit (159). MDSCs are immature myeloid cells with
strong immunosuppressive activity, which can trigger the
immunosuppressive tumor microenvironment and promote tumor growth
(160). In addition, TLR2 signaling
also enhanced the immunosuppressive ability of MDSCs on NK cells
via a paracrine IL-6/JAK1/2-dependent mechanism. Therefore, this
TRF2-dependent HSPG biosynthesis gene regulatory network promoted
the recruitment and function of MDSCs, inhibited the recruitment
and cytotoxicity of NK cells and immune lymphocytes, and was
negatively associated with the overall survival (OS) time in
patients with gastric cancer, as it ultimately leads to tumor
progression and metastasis (159).
Targeting TRF2 and tumor therapy
TRF2 is considered as a potential new target for
cancer treatment. Inhibition of TRF2 could activate ATM-dependent
DDR pathways and induce cell apoptosis or senescence (161). When short interfering RNA was used
to knockdown the expression level of TRF2 in colorectal cancer and
renal epithelial renal cell carcinoma cell lines, tumor cell
proliferation and clonal formation were notably inhibited, and cell
cycle arrest and apoptosis were markedly induced (125,162).
In the xenotransplantation experiment of intracranial tumors, GSCs
infected by lentivirus shTRF2 and empty vector were stereotaxically
injected into the cerebral cortex of BALB/c nude mice, which lack a
thymus. The results demonstrated that the life span of mice in the
experimental group was significantly prolonged compared with that
in the control group (157). This
finding revealed that reducing the expression level of TRF2 may
notably improve the survival rate of nude mice in a glioblastoma
model. The aforementioned results all indicate that TRF2 may be a
new antitumor treatment target. Furthermore, previous studies
(163,164) suggest that, compared with that in
the previous strategy of inhibiting telomerase to treat tumors,
tumor therapy targeting TRF2 may be more effective. The main
reasons are as follows: First, telomerase inhibitors were not
effective on tumor cells that rely on the ALT mechanism; second, in
tumors exhibiting increased TRF2 expression, the therapeutic effect
of telomerase inhibitors was not notable (163); reducing the expression level of TRF2
shortened the 3′ overhangs of telomeres, leading to chromosomal
fusion of tumor cells and premature senescence or death of tumor
cells. Decreasing the expression level of TRF2 could activate
phosphorylated ATM, activating the tumor suppressor gene p53, which
is not only an effector molecule downstream of the telomere damage
signal, but also one of the main senescence promoters. P53
expresses the E3 ubiquitin ligase, Siah1, and targets TRF2 for
ubiquitination and proteasomal degradation (164). TRF2 knockdown further promoted ATM
activation; therefore, TRF2/ATM/p53/Siah1 formed a positive
feedback loop, which could simultaneously promote telomere
decapsulation, induce the p53-mediated cellular senescence signal
transduction pathway, and enhance tumor cell suppression (164). Therefore, tumor therapy targeting
TRF2 has broad application prospects.
Radio- and chemotherapy play an important role in
the treatment of malignant tumors; however, tumor drug resistance
remains one of the main obstacles in clinical treatment. An
increasing number of studies have found that TRF2 was associated
with tumor drug resistance. In radioresistant A549R
(telomerase-positive) and U2OSR (ALT mechanism) cells, TRF2
expression was found to be notably increased, while decreasing the
expression level of TRF2 notably improved the radiosensitivity of
tumor cells (165–167). It has been confirmed that there was
a notable negative correlation between telomere length and
radiosensitivity (166,167). Knockout of TRF2 may lead to telomere
shortening and inhibition of telomerase activity, leading to
radiosensitization and shortening of telomeres in A549 and U2OS
cells, suggesting that TRF2 may regulate radiosensitization via
telomere length and DDR. Orun et al (168) also found the same results while
investigating immortalized mesenchymal stem cells, suggesting that
TRF2 may be a potential target of radiotherapy for tumors. Ning
et al (169,170) found that the expression level of
TRF2 in the vincristine-resistant gastric cancer cell line,
SGC790l/VCR, and the adriamycin-resistant gastric cancer cell line,
SGC7901/ADR, was notably higher compared with that in parental
cells. Inhibition of TRF2 expression could notably enhance the
sensitivity of drug-resistant cells to cisplatin, adriamycin and
etoposide (170). Furthermore, it
was found that the expression of ATM, γ-H2AX and p53 in
drug-resistant cells was notably lower compared with that in
SGC7901 and, after using shRNA to inhibit the expression level of
TRF2, the expression levels of ATM, γ-H2AX and p53 were notably
upregulated, suggesting that TRF2 may promote tumor drug resistance
by inhibiting the expression of ATM and its downstream target
molecules (169,170). Benhamou et al (171) also found that inhibiting the
expression level of TRF2 could notably enhance the sensitivity of
oral squamous cell carcinoma cells to EGFR-targeting
chemotherapeutics, such as cetuximab and erlotinib, which may
become an important development and progress in cancer treatment.
It has also been found that, in addition to TRF2 inhibition
activating the ATM-dependent DDR pathways and inducing cell
apoptosis or senescence, it can also inhibit p38MAPK
phosphorylation, block tumor pathway signal transduction and
increase chemotherapeutic drug sensitivity (172).
In mice with tumor overexpressing TRF2, a decrease
in tumor volume and an increase in OS time were observed following
treatment with fluorouracil (5-FU), indicating that TRF2
overexpression may increase the treatment efficiency of 5-FU
(159). This was consistent with the
MDSC knockdown results, and the enhanced response to 5-FU may be
associated with enhanced elimination of MDSCs, allowing the immune
system to become reactivated (159).
Therefore, the expression level of TRF2 may be an interesting
surrogate indicator of response to chemotherapy (159). The use of 5-FU treatment in gastric
cancer, gemcitabine treatment in ovarian cancer, and
paclitaxel-based chemotherapy in lung cancer (all 3 drugs target
MDSCs) could reduce the negative effect of TRF2 overexpression on
OS time (173–175). This indicates that the efficiency of
chemotherapy is improved in patients with cancer and TRF2
overexpression.
A previous study (176) showed that the small molecule,
curcusone C may bind to TRF2 via its DNA-binding site, thereby
hindering its interaction with telomere DNA, activating the DDR,
inhibiting tumor cell proliferation, causing cell cycle arrest and
tumor cell apoptosis. Curcusone C may become a promising lead
compound for the treatment of cancer with elevated TRF2. The
anticancer activity of other terpenoids may be attributed to their
effect on telomere protective protein. Chromosome analysis revealed
that As2O3 downregulated the expression level of TRF2, contributing
to end-to-end fusion and apoptosis of leukemia cells. Inhibition of
TRF2 rendered leukemia cells more sensitive to As2O3, suggesting
that TRF2 may represent a promising target for novel leukemia
therapies (176).
In summary, maintaining the function of TRF2 is
crucial for the control of pathological tumor cell behaviors, such
as proliferation, invasion and migration. Overexpression or
knockdown of TRF2 may cause CIN, affecting genome expression and
cell survival. Regulating the function of TRF2, using artificial
methods, to determine the prognosis of tumors, and its potential
value in tumor treatment, require further in-depth investigations.
Therefore, the research and development of TRF2 functional
knockdown or function-regulating drugs has broad prospects, and it
will likely become one of the important research directions in
tumor diagnosis and treatment in the future.
Conclusion
Accumulating evidence indicates that the telomere
and non-telomere-related functions of the TRF2 protein play a key
role in the regulation of cell lifespan, chromosome stability, DNA
repair or replication, and the extracellular microenvironment
(5–8).
TRF2 links multiple functions or signaling pathways, which cells
rely on for survival and reproduction, performs systemic
regulation, enhances cell environment adaptation and survival
ability, assists with the elimination of senescent mutant cells,
and maintains the stability of genetic material (49,95,131,143).
When replicative aging or other pathological factors (such as
ionizing radiation) cause TRF2 dysfunction, cell physiological
balance is severely disrupted, telomeres lose protein protection,
telomere caps are eliminated, DDR is triggered, cell cycle is
arrested, DNA is abnormally repaired, and end-to-end fusions occur.
At this time, the chromosome is already unstable and is easily
mutated (96–99). Due to cell checkpoint blockade, most
cells undergo senescent apoptosis during this period to maintain
the genetic stability of the body. However, a small number of cells
may induce mutations, such as in p53 protein, which is common in
most malignant tumors or abnormal expression of TRF2, disrupting
the DDR cascade and, by extending the mitotic arrest cycle,
bypassing the DDR-induced apoptosis mechanism and entering the
telomere crisis period (100).
During the telomere crisis, chromosomes are recombined via the BFB
cycle DNA to generate large-scale mutant cells, referred to as
precancerous cells. Finally, at the end of this cycle, telomerase
expression or the ALT mechanism is activated to extend local
telomeres, and gradually stabilize chromosomes under the protection
of TRF2, forming immortal cells (6,101–103).
This may be the stagewise mechanism underlying the transformation
of normal human cells to malignant cells. Due to the uncertainty
and randomness of DNA recombination in the BFB cycle, the mutation
itself is heterogeneous and can generate the same immortal cells
with heterogeneous characteristics, which may be an important
origin of tumor heterogeneity (104–106).
Subsequently, tumor cells have a telomere lengthening mechanism,
due to the abnormal proliferation; however, the telomere remains
relatively short, and the chromosome is only in a relatively stable
state, which can produce subsequent gene recombinations. This may
be the basis of mutations and typing of various subtypes of tumor
cells (104–106). After the formation of immortalized
tumor cells, the increase in TRF2 expression levels has an effect
on the structure of the glycocalyx outside the tumor cell membrane
and constitutes a microenvironment conducive to tumor growth. In
this microenvironment, chemotactic aggregation of immunosuppressive
MDSCs, and paracrine immunosuppressive factors, inhibit the tumor
killing activity of NK cells and T lymphocytes (8,157–159).
Furthermore, TRF2 acts synergistically with multiple signaling
pathways in tumor cell proliferation and migration to maintain its
own intracellular levels and promote tumor development (131,138,141,142,144,146,147).
The function of TRF2 is also indispensable for maintaining the
stemness characteristics of CSCs. TRF2 interacts with REST, KLF4
and other proteins (such asOct4 and Sox2) to increase the activity
of tumor cell stemness-related transcription factors, promote tumor
stem cell division and proliferation, drug resistance, EMT,
invasion and metastasis, which have a major effect on tumor
prognosis (150,154,156,157).
With the proliferation of tumor cells and the continuous
development of the tumor, via the effect of angiogenesis-related
factors or receptors, such as VEGF-A and PDGFRβ, TRF2 can promote
the formation of nourishing tumor blood vessels, which facilitates
the nutrient uptake and metabolism of tumor cells, further
enhancing tumor development and contributing to a poor prognosis
(7,136,137).
Therefore, the targeted treatment of TRF2 is
important and has potential. Inhibition of TRF2 disrupts several
pathways in which tumor cells depend for survival: It improves the
immunosuppressive tumor microenvironment, promotes tumor cell
apoptosis and CSC differentiation, inhibits the activity of
telomerase and the ALT mechanism, and enhances the sensitivity of
cancer cells to radiotherapy and chemotherapy (157,160–164,166).
Under the action of the aforementioned multiple anticancer factors,
it is difficult for tumor cells to continue to proliferate or
survive; therefore, the efficiency of tumor treatment is increased.
After TRF2 expression level is knocked down, telomerase and ALT
functions are also simultaneously inhibited, and the
transcriptional activity of CSC stemness-related factors is
blocked, which is conducive to the differentiation of CSCs and the
loss of immortalization ability (102,103,112,113,150).
This can effectively inhibit the accumulation of malignant cell
mutant genes in vivo, reduce the invasion and metastasis of
tumor cells under the stress of treatment, and effectively remove
mutated cells. However, with respect to safety, TRF2-targeted
treatment may cause senescence and apoptosis of normal cells.
Furthermore, specific adverse reactions require further evaluation.
The specific targeting of tumor cell TRF2 is an issue, which
requires resolving in TRF2 treatment research; however, the
therapeutic prospects of this new target are worthy of further
investigation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Author's contributions
Both authors (ZW and XW) made substantial
contributions to the collection, sorting, image processing,
editing, writing, proofreading and revision of the manuscript, and
approved the final version. Data sharing is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Martincorena I, Fowler JC, Wabik A, Lawson
ARJ, Abascal F, Hall MWJ, Cagan A, Murai K, Mahbubani K, Stratton
MR, et al: Somatic mutant clones colonize the human esophagus with
age. Science. 362:911–917. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yokoyama A, Kakiuchi N, Yoshizato T,
Nannya Y, Suzuki H, Takeuchi Y, Shiozawa Y, Sato Y, Aoki K, Kim SK,
et al: Age-related remodelling of oesophageal epithelia by mutated
cancer drivers. Nature. 565:312–317. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goerttler K, Loehrke H, Schweizer J and
Hesse B: Two-stage skin carcinogenesis by systemic initiation of
pregnant mice with 7,12-dimethylbenz(a)anthracene during gestation
days 6–20 and postnatal promotion of the F 1-generation with the
phorbol ester 12-tetradecanoylphorbol-13-acetate. J Cancer Res Clin
Oncol. 98:267–275. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goerttler K, Loehrke H, Hesse B, Milz A
and Schweizer J: Diaplacental initiation of NMRI mice with
7,12-dimethylbenz[a]anthracene during gestation days 6–20 and
postnatal treatment of the F1-generation with the phorbol ester
12-O-tetradecanoylphorbol-13-acetate: Tumor incidence in organs
other than the skin. Carcinogenesis. 2:1087–1094. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fairall L, Chapman L, Moss H, de Lange T
and Rhodes D: Structure of the TRFH dimerization domain of the
human telomeric proteins TRF1 and TRF2. Mol Cell. 8:351–361. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mondello C, Smirnova A and Giulotto E:
Gene amplification, radiation sensitivity and DNA double-strand
breaks. Mutat Res. 704:29–37. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
El Maï M, Wagner KD, Michiels JF,
Ambrosetti D, Borderie A, Destree S, Renault V, Djerbi N,
Giraud-Panis MJ, Gilson E and Wagner N: The telomeric protein TRF2
regulates angiogenesis by binding and activating the PDGFRβ
promoter. Cell Rep. 9:1047–1060. 2014. View Article : Google Scholar
|
|
8
|
Biroccio A, Cherfils-Vicini J, Augereau A,
Pinte S, Bauwens S, Ye J, Simonet T, Horard B, Jamet K, Cervera L,
et al: TRF2 inhibits a cell-extrinsic pathway through which natural
killer cells eliminate cancer cells. Nat Cell Biol. 15:818–828.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van Steensel B, Smogorzewska A and de
Lange T: TRF2 protects human telomeres from end-to-end fusions.
Cell. 92:401–413. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Lange T: Shelterin: The protein complex
that shapes and safeguards human telomeres. Genes Dev.
19:2100–2110. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smith EM, Pendlebury DF and Nandakumar J:
Structural biology of telomeres and telomerase. Cell Mol Life Sci.
77:61–79. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deng Z, Norseen J, Wiedmer A, Riethman H
and Lieberman PM: TERRA RNA binding to TRF2 facilitates
heterochromatin formation and ORC recruitment at telomeres. Mol
Cell. 35:403–413. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Necasová I, Janoušková E, Klumpler T and
Hofr C: Basic domain of telomere guardian TRF2 reduces D-loop
unwinding whereas Rap1 restores it. Nucleic Acids Res.
45:12170–12180. 2017. View Article : Google Scholar
|
|
14
|
O'Connor MS, Safari A, Xin H, Liu D and
Songyang Z: A critical role for TPP1 and TIN2 interaction in
high-order telomeric complex assembly. Proc Natl Acad Sci USA.
103:11874–11879. 2006. View Article : Google Scholar
|
|
15
|
Xin H, Liu D, Wan M, Safari A, Kim H, Sun
W, O'Connor MS and Songyang Z: TPP1 is a homologue of ciliate
TEBP-beta and interacts with POT1 to recruit telomerase. Nature.
445:559–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
van Overbeek M and de Lange T: Apollo, an
Artemis-related nuclease, interacts with TRF2 and protects human
telomeres in S phase. Curr Biol. 16:1295–1302. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lenain C, Bauwens S, Amiard S, Brunori M,
Giraud-Panis MJ and Gilson E: The Apollo 5′exonuclease functions
together with TRF2 to protect telomeres from DNA repair. Curr Biol.
16:1303–1310. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim H, Lee OH, Xin H, Chen LY, Qin J, Chae
HK, Lin SY, Safari A, Liu D and Songyang Z: TRF2 functions as a
protein hub and regulates telomere maintenance by recognizing
specific peptide motifs. Nat Struct Mol Biol. 16:372–379. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Benarroch-Popivker D, Pisano S,
Mendez-Bermudez A, Lototska L, Kaur P, Bauwens S, Djerbi N, Latrick
CM, Fraisier V, Pei B, et al: TRF2-mediated control of telomere DNA
topology as a mechanism for chromosome-end protection. Mol Cell.
61:274–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Giraud-Panis MJ, Pisano S,
Benarroch-Popivker D, Pei B, Le Du MH and Gilson E: One identity or
more for telomeres? Front Oncol. 3:482013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Broccoli D, Smogorzewska A, Chong L and de
Lange T: Human telomeres contain two distinct Myb-related proteins,
TRF1 and TRF2. Nat Genet. 17:231–235. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Baker AM, Fu Q, Hayward W, Lindsay SM and
Fletcher TM: The Myb/SANT domain of the telomere-binding protein
TRF2 alters chromatin structure. Nucleic Acids Res. 37:5019–5031.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bilaud T, Brun C, Ancelin K, Koering CE,
Laroche T and Gilson E: Telomeric localization of TRF2, a novel
human telobox protein. Nat Genet. 17:236–239. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Denchi EL and de Lange T: Protection of
telomeres through independent control of ATM and ATR by TRF2 and
POT1. Nature. 448:1068–1071. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gilson E and Géli V: How telomeres are
replicated. Nat Rev Mol Cell Biol. 8:825–838. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ye J, Lenain C, Bauwens S, Rizzo A,
Saint-Léger A, Poulet A, Benarroch D, Magdinier F, Morere J, Amiard
S, et al: TRF2 and apollo cooperate with topoisomerase 2alpha to
protect human telomeres from replicative damage. Cell. 142:230–242.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Greider CW: Telomeres do D-loop-T-loop.
Cell. 97:419–422. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Griffith JD, Comeau L, Rosenfield S,
Stansel RM, Bianchi A, Moss H and de Lange T: Mammalian telomeres
end in a large duplex loop. Cell. 97:503–514. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Timashev LA and De Lange T:
Characterization of t-loop formation by TRF2. Nucleus. 11:164–177.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feuerhahn S, Chen LY, Luke B and Porro A:
No DDRama at chromosome ends: TRF2 takes centre stage. Trends
Biochem Sci. 40:275–285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmutz I, Timashev L, Xie W, Patel DJ and
de Lange T: TRF2 binds branched DNA to safeguard telomere
integrity. Nat Struct Mol Biol. 24:734–742. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sarek G, Kotsantis P, Ruis P, Van Ly D,
Margalef P, Borel V, Zheng XF, Flynn HR, Snijders AP, Chowdhury D,
et al: CDK phosphorylation of TRF2 controls t-loop dynamics during
the cell cycle. Nature. 575:523–527. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sarek G, Vannier JB, Panier S, Petrini JHJ
and Boulton SJ: TRF2 Recruits RTEL1 to Telomeres in S Phase to
Promote T-Loop Unwinding. Mol Cell. 61:788–789. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bower BD and Griffith JD: TRF1 and TRF2
differentially modulate Rad51-mediated telomeric and nontelomeric
displacement loop formation in vitro. Biochemistry. 53:5485–5495.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
d'Adda di Fagagna F, Reaper PM,
Clay-Farrace L, Fiegler H, Carr P, Von Zglinicki T, Saretzki G,
Carter NP and Jackson SP: A DNA damage checkpoint response in
telomere-initiated senescence. Nature. 426:194–198. 2003.
View Article : Google Scholar
|
|
36
|
Takai H, Smogorzewska A and de Lange T:
DNA damage foci at dysfunctional telomeres. Curr Biol.
13:1549–1556. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Okamoto K, Bartocci C, Ouzounov I,
Diedrich JK, Yates JR III and Denchi EL: A two-step mechanism for
TRF2-mediated chromosome-end protection. Nature. 494:502–505. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Frescas D and de Lange T: TRF2-tethered
TIN2 can mediate telomere protection by TPP1/POT1. Mol Cell Biol.
34:1349–1362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Takai KK, Kibe T, Donigian JR, Frescas D
and de Lange T: Telomere protection by TPP1/POT1 requires tethering
to TIN2. Mol Cell. 67:1622017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fuchs E: The tortoise and the hair:
Slow-cycling cells in the stem cell race. Cell. 137:811–819. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Opresko PL, von Kobbe C, Laine JP,
Harrigan J, Hickson ID and Bohr VA: Telomere-binding protein TRF2
binds to and stimulates the Werner and Bloom syndrome helicases. J
Biol Chem. 277:41110–41119. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Stavropoulos DJ, Bradshaw PS, Li X, Pasic
I, Truong K, Ikura M, Ungrin M and Meyn MS: The Bloom syndrome
helicase BLM interacts with TRF2 in ALT cells and promotes
telomeric DNA synthesis. Hum Mol Genet. 11:3135–3144. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Rothkamm K, Krüger I, Thompson LH and
Löbrich M: Pathways of DNA double-strand break repair during the
mammalian cell cycle. Mol Cell Biol. 23:5706–5715. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mladenov E and Iliakis G: Induction and
repair of DNA double strand breaks: The increasing spectrum of
non-homologous end joining pathways. Mutat Res. 711:61–72. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ribes-Zamora A, Indiviglio SM, Mihalek I,
Williams CL and Bertuch AA: TRF2 interaction with Ku
heterotetramerization interface gives insight into c-NHEJ
prevention at human telomeres. Cell Rep. 5:194–206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dimitrova N, Chen YC, Spector DL and de
Lange T: 53BP1 promotes non-homologous end joining of telomeres by
increasing chromatin mobility. Nature. 456:524–528. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mirman Z, Lottersberger F, Takai H, Kibe
T, Gong Y, Takai K, Bianchi A, Zimmermann M, Durocher D and de
Lange T: 53BP1-RIF1-shieldin counteracts DSB resection through CST-
and Polα-dependent fill-in. Nature. 560:112–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Timashev LA, Babcock H, Zhuang X and de
Lange T: The DDR at telomeres lacking intact shelterin does not
require substantial chromatin decompaction. Genes Dev. 31:578–589.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Blasco MA: Telomeres and human disease:
Ageing, cancer and beyond. Nat Rev Genet. 6:611–622. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sfeir A and de Lange T: Removal of
shelterin reveals the telomere end-protection problem. Science.
336:593–597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Celli GB, Denchi EL and de Lange T: Ku70
stimulates fusion of dysfunctional telomeres yet protects
chromosome ends from homologous recombination. Nat Cell Biol.
8:885–890. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kibe T, Osawa GA, Keegan CE and de Lange
T: Telomere protection by TPP1 is mediated by POT1a and POT1b. Mol
Cell Biol. 30:1059–1066. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shamanna RA, Lu H, de Freitas JK, Tian J,
Croteau DL and Bohr VA: WRN regulates pathway choice between
classical and alternative non-homologous end joining. Nat Commun.
7:137852016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fallet E, Jolivet P, Soudet J, Lisby M,
Gilson E and Teixeira MT: Length-dependent processing of telomeres
in the absence of telomerase. Nucleic Acids Res. 42:3648–3665.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu L, Multani AS, He H, Cosme-Blanco W,
Deng Y, Deng JM, Bachilo O, Pathak S, Tahara H, Bailey SM, et al:
Pot1 deficiency initiates DNA damage checkpoint activation and
aberrant homologous recombination at telomeres. Cell. 126:49–62.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rai R, Chen Y, Lei M and Chang S:
TRF2-RAP1 is required to protect telomeres from engaging in
homologous recombination-mediated deletions and fusions. Nat
Commun. 7:108812016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
González-Prieto R, Cuijpers SA,
Luijsterburg MS, van Attikum H and Vertegaal AC: SUMOylation and
PARylation cooperate to recruit and stabilize SLX4 at DNA damage
sites. EMBO Rep. 16:512–519. 2015. View Article : Google Scholar
|
|
58
|
Capper R, Britt-Compton B, Tankimanova M,
Rowson J, Letsolo B, Man S, Haughton M and Baird DM: The nature of
telomere fusion and a definition of the critical telomere length in
human cells. Genes Dev. 21:2495–2508. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Karlseder J, Smogorzewska A and de Lange
T: Senescence induced by altered telomere state, not telomere loss.
Science. 295:2446–2449. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Smogorzewska A, van Steensel B, Bianchi A,
Oelmann S, Schaefer MR, Schnapp G and de Lange T: Control of human
telomere length by TRF1 and TRF2. Mol Cell Biol. 20:1659–1668.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Muñoz P, Blanco R and Blasco MA: Role of
the TRF2 telomeric protein in cancer and ageing. Cell Cycle.
5:718–721. 2006. View Article : Google Scholar
|
|
62
|
Saint-Léger A, Koelblen M, Civitelli L,
Bah A, Djerbi N, Giraud-Panis MJ, Londoño-Vallejo A, Ascenzioni F
and Gilson E: The basic N-terminal domain of TRF2 limits
recombination endonuclease action at human telomeres. Cell Cycle.
13:2469–2474. 2014. View Article : Google Scholar
|
|
63
|
Wilson JS, Tejera AM, Castor D, Toth R,
Blasco MA and Rouse J: Localization-dependent and -independent
roles of SLX4 in regulating telomeres. Cell Rep. 4:853–860. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wu Y, Zacal NJ, Rainbow AJ and Zhu XD: XPF
with mutations in its conserved nuclease domain is defective in DNA
repair but functions in TRF2-mediated telomere shortening. DNA
Repair (Amst). 6:157–166. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wu Y, Mitchell TR and Zhu XD: Human XPF
controls TRF2 and telomere length maintenance through distinctive
mechanisms. Mech Ageing Dev. 129:602–610. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Menendez JA, Rubio MA, Campisi J and Lupu
R: Heregulin, a new regulator of telomere length in human cells.
Oncotarget. 6:39422–39436. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Menendez JA, Benboudjema L, Vellon L,
Rubio MA, Espinoza I, Campisi J and Lupu R: Heregulin, a new
interactor of the telosome/shelterin complex in human telomeres.
Oncotarget. 6:39408–39421. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Baur JA, Zou Y, Shay JW and Wright WE:
Telomere position effect in human cells. Science. 292:2075–2077.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kim W, Ludlow AT, Min J, Robin JD, Stadler
G, Mender I, Lai TP, Zhang N, Wright WE and Shay JW: Regulation of
the human telomerase gene TERT by telomere position effect-over
long distances (TPE-OLD): Implications for aging and cancer. PLoS
Biol. 14:e20000162016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Mukherjee AK, Sharma S, Bagri S, Kutum R,
Kumar P, Hussain A, Singh P, Saha D, Kar A, Dash D and Chowdhury S:
Telomere repeat-binding factor 2 binds extensively to
extra-telomeric G-quadruplexes and regulates the epigenetic status
of several gene promoters. J Biol Chem. 294:17709–17722. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bradshaw PS, Stavropoulos DJ and Meyn MS:
Human telomeric protein TRF2 associates with genomic double-strand
breaks as an early response to DNA damage. Nat Genet. 37:193–197.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hussain T, Saha D, Purohit G, Kar A,
Kishore Mukherjee A, Sharma S, Sengupta S, Dhapola P, Maji B,
Vedagopuram S, et al: Transcription regulation of CDKN1A
(p21/CIP1/WAF1) by TRF2 is epigenetically controlled through the
REST repressor complex. Sci Rep. 7:115412017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Purohit G, Mukherjee AK, Sharma S and
Chowdhury S: Extratelomeric binding of the telomere binding protein
TRF2 at the PCGF3 promoter is G-Quadruplex Motif-dependent.
Biochemistry. 57:2317–2324. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Williamson JR, Raghuraman MK and Cech TR:
Monovalent cation-induced structure of telomeric DNA: The G-quartet
model. Cell. 59:871–880. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Phan AT: Human telomeric G-quadruplex:
Structures of DNA and RNA sequences. FEBS J. 277:1107–1117. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Muniyappa K and Kironmai KM: Telomere
structure, replication and length maintenance. Crit Rev Biochem Mol
Biol. 33:297–336. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Luu KN, Phan AT, Kuryavyi V, Lacroix L and
Patel DJ: Structure of the human telomere in K+ solution: An
intramolecular (3 + 1) G-quadruplex scaffold. J Am Chem Soc.
128:9963–9970. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Brázda V, Hároníková L, Liao JC and Fojta
M: DNA and RNA quadruplex-binding proteins. Int J Mol Sci.
15:17493–17517. 2014. View Article : Google Scholar
|
|
79
|
Pedroso IM, Hayward W and Fletcher TM: The
effect of the TRF2 N-terminal and TRFH regions on telomeric
G-quadruplex structures. Nucleic Acids Res. 37:1541–1554. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Rawal P, Kummarasetti VB, Ravindran J,
Kumar N, Halder K, Sharma R, Mukerji M, Das SK and Chowdhury S:
Genome-wide prediction of G4 DNA as regulatory motifs: Role in
Escherichia coli global regulation. Genome Res. 16:644–655. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Sengupta A, Roy SS and Chowdhury S:
Non-duplex G-Quadruplex DNA Structure: A developing story from
predicted sequences to DNA structure-dependent epigenetics and
beyond. Acc Chem Res. 54:46–56. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Killela PJ, Pirozzi CJ, Healy P, Reitman
ZJ, Lipp E, Rasheed BA, Yang R, Diplas BH, Wang Z, Greer PK, et al:
Mutations in IDH1, IDH2, and in the TERT promoter define clinically
distinct subgroups of adult malignant gliomas. Oncotarget.
5:1515–1525. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Heidenreich B, Nagore E, Rachakonda PS,
Garcia-Casado Z, Requena C, Traves V, Becker J, Soufir N, Hemminki
K and Kumar R: Telomerase reverse transcriptase promoter mutations
in primary cutaneous melanoma. Nat Commun. 5:34012014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Pinyol R, Tovar V and Llovet JM: TERT
promoter mutations: Gatekeeper and driver of hepatocellular
carcinoma. J Hepatol. 61:685–687. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Rachakonda PS, Hosen I, de Verdier PJ,
Fallah M, Heidenreich B, Ryk C, Wiklund NP, Steineck G, Schadendorf
D, Hemminki K and Kumar R: TERT promoter mutations in bladder
cancer affect patient survival and disease recurrence through
modification by a common polymorphism. Proc Natl Acad Sci USA.
110:17426–17431. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Thakur RK, Kumar P, Halder K, Verma A, Kar
A, Parent JL, Basundra R, Kumar A and Chowdhury S: Metastases
suppressor NM23-H2 interaction with G-quadruplex DNA within c-MYC
promoter nuclease hypersensitive element induces c-MYC expression.
Nucleic Acids Res. 37:172–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Paramasivam M, Membrino A, Cogoi S, Fukuda
H, Nakagama H and Xodo LE: Protein hnRNP A1 and its derivative Up1
unfold quadruplex DNA in the human KRAS promoter: Implications for
transcription. Nucleic Acids Res. 37:2841–2853. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Cogoi S, Paramasivam M, Membrino A,
Yokoyama KK and Xodo LE: The KRAS promoter responds to
Myc-associated zinc finger and poly(ADP-ribose) polymerase 1
proteins, which recognize a critical quadruplex-forming GA-element.
J Biol Chem. 285:22003–22016. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yadav VK, Abraham JK, Mani P, Kulshrestha
R and Chowdhury S: QuadBase: Genome-wide database of G4
DNA-occurrence and conservation in human, chimpanzee, mouse and rat
promoters and 146 microbes. Nucleic Acids Res. 36:D381–D385. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Mukherjee AK, Sharma S, Sengupta S, Saha
D, Kumar P, Hussain T, Srivastava V, Roy SD, Shay JW and Chowdhury
S: Telomere length-dependent transcription and epigenetic
modifications in promoters remote from telomere ends. PLoS Genet.
14:e10077822018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Shay JW and Wright WE: Telomeres and
telomerase: Three decades of progress. Nat Rev Genet. 20:299–309.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Rajagopalan H and Lengauer C: CIN-ful
cancers. Cancer Chemother Pharmacol. 54 (Suppl 1):S65–S68.
2004.PubMed/NCBI
|
|
93
|
Tlsty TD: Genomic instability and its role
in neoplasia. Curr Top Microbiol Immunol. 221:37–46.
1997.PubMed/NCBI
|
|
94
|
Lengauer C, Kinzler KW and Vogelstein B:
Genetic instabilities in human cancers. Nature. 396:643–649. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Palm W and de Lange T: How shelterin
protects mammalian telomeres. Annu Rev Genet. 42:301–334. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wright WE, Pereira-Smith OM and Shay JW:
Reversible cellular senescence: Implications for immortalization of
normal human diploid fibroblasts. Mol Cell Biol. 9:3088–3092. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
O'Hagan RC, Chang S, Maser RS, Mohan R,
Artandi SE, Chin L and DePinho RA: Telomere dysfunction provokes
regional amplification and deletion in cancer genomes. Cancer Cell.
2:149–155. 2002. View Article : Google Scholar
|
|
98
|
Fumagalli M, Rossiello F, Clerici M,
Barozzi S, Cittaro D, Kaplunov JM, Bucci G, Dobreva M, Matti V,
Beausejour CM, et al: Telomeric DNA damage is irreparable and
causes persistent DNA-damage-response activation. Nat Cell Biol.
14:355–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Feldser DM and Greider CW: Short telomeres
limit tumor progression in vivo by inducing senescence. Cancer
Cell. 11:461–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Counter CM, Avilion AA, LeFeuvre CE,
Stewart NG, Greider CW, Harley CB and Bacchetti S: Telomere
shortening associated with chromosome instability is arrested in
immortal cells which express telomerase activity. EMBO J.
11:1921–1929. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Hayashi MT, Cesare AJ, Rivera T and
Karlseder J: Cell death during crisis is mediated by mitotic
telomere deprotection. Nature. 522:492–496. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Sabatier L, Ricoul M, Pottier G and
Murnane JP: The loss of a single telomere can result in instability
of multiple chromosomes in a human tumor cell line. Mol Cancer Res.
3:139–150. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Bailey SM and Murnane JP: Telomeres,
chromosome instability and cancer. Nucleic Acids Res. 34:2408–2417.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Maciejowski J and de Lange T: Telomeres in
cancer: Tumour suppression and genome instability. Nat Rev Mol Cell
Biol. 18:175–186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Martínez P and Blasco MA: Telomere-driven
diseases and telomere-targeting therapies. J Cell Biol.
216:875–887. 2017. View Article : Google Scholar
|
|
106
|
Murnane JP and Sabatier L: Chromosome
rearrangements resulting from telomere dysfunction and their role
in cancer. Bioessays. 26:1164–1174. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Giraud-Panis MJ, Pisano S, Poulet A, Le Du
MH and Gilson E: Structural identity of telomeric complexes. FEBS
Lett. 584:3785–3799. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Salhab M, Jiang WG, Newbold RF and Mokbel
K: The expression of gene transcripts of telomere-associated genes
in human breast cancer: Correlation with clinico-pathological
parameters and clinical outcome. Breast Cancer Res Treat.
109:35–46. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Pascua I, Fernández-Marcelo T,
Sánchez-Pernaute A, de Juan C, Head J, Torres-García AJ and Iniesta
P: Prognostic value of telomere function in gastric cancers with
and without microsatellite instability. Eur J Gastroenterol
Hepatol. 27:162–169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Yamada K, Yagihashi A, Yamada M, Asanuma
K, Moriai R, Kobayashi D, Tsuji N and Watanabe N: Decreased gene
expression for telomeric-repeat binding factors and TIN2 in
malignant hematopoietic cells. Anticancer Res. 22:1315–1320.
2002.PubMed/NCBI
|
|
111
|
Su CH, Cheng C, Tzeng TY, Lin IH and Hsu
MT: An H2A Histone Isotype, H2ac, associates with telomere and
maintains telomere integrity. PLoS One. 11:e01563782016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Bojovic B, Ho HY, Wu J and Crowe DL: Stem
cell expansion during carcinogenesis in stem cell-depleted
conditional telomeric repeat factor 2 null mutant mice. Oncogene.
32:5156–5166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Lagunas AM, Wu J and Crowe DL: Telomere
DNA damage signaling regulates cancer stem cell evolution,
epithelial mesenchymal transition, and metastasis. Oncotarget.
8:80139–80155. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Rossi DJ, Jamieson CH and Weissman IL:
Stems cells and the pathways to aging and cancer. Cell.
132:681–696. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Härle-Bachor C and Boukamp P: Telomerase
activity in the regenerative basal layer of the epidermis inhuman
skin and in immortal and carcinoma-derived skin keratinocytes. Proc
Natl Acad Sci USA. 93:6476–6481. 1996. View Article : Google Scholar
|
|
117
|
González-Suárez E, Samper E, Ramírez A,
Flores JM, Martín-Caballero J, Jorcano JL and Blasco MA: Increased
epidermal tumors and increased skin wound healing in transgenic
mice overexpressing the catalytic subunit of telomerase, mTERT, in
basal keratinocytes. EMBO J. 20:2619–2630. 2001. View Article : Google Scholar
|
|
118
|
Blanco R, Muñoz P, Flores JM, Klatt P and
Blasco MA: Telomerase abrogation dramatically accelerates
TRF2-induced epithelial carcinogenesis. Genes Dev. 21:206–220.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Matsutani N, Yokozaki H and Tahara E,
Tahara H, Kuniyasu H, Haruma K, Chayama K, Yasui W and Tahara E:
Expression of telomeric repeat binding factor 1 and 2 and
TRF1-interacting nuclear protein 2 in human gastric carcinomas. Int
J Oncol. 19:507–512. 2001.PubMed/NCBI
|
|
120
|
Oh BK, Kim YJ, Park C and Park YN:
Up-regulation of telomere-binding proteins, TRF1, TRF2, and TIN2 is
related to telomere shortening during human multistep
hepatocarcinogenesis. Am J Pathol. 166:73–80. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Xu L and Blackburn EH: Human cancer cells
harbor T-stumps, a distinct class of extremely short telomeres. Mol
Cell. 28:315–327. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Muñoz P, Blanco R, Flores JM and Blasco
MA: XPF nuclease-dependent telomere loss and increased DNA damage
in mice overexpressing TRF2 result in premature aging and cancer.
Nat Genet. 37:1063–1071. 2005. View
Article : Google Scholar
|
|
123
|
Nera B, Huang HS, Lai T and Xu L: Elevated
levels of TRF2 induce telomeric ultrafine anaphase bridges and
rapid telomere deletions. Nat Commun. 6:101322015. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Rai R, Zheng H, He H, Luo Y, Multani A,
Carpenter PB and Chang S: The function of classical and alternative
non-homologous end-joining pathways in the fusion of dysfunctional
telomeres. EMBO J. 29:2598–2610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Dong W, Shen R, Wang Q, Gao Y, Qi X, Jiang
H, Yao J, Lin X, Wu Y and Wang L: Sp1 upregulates expression of
TRF2 and TRF2 inhibition reduces tumorigenesis in human colorectal
carcinoma cells. Cancer Biol Ther. 8:2166–2174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Raynaud CM, Jang SJ, Nuciforo P,
Lantuejoul S, Brambilla E, Mounier N, Olaussen KA, André F, Morat
L, Sabatier L and Soria JC: Telomere shortening is correlated with
the DNA damage response and telomeric protein down-regulation in
colorectal preneoplastic lesions. Ann Oncol. 19:1875–1881. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Brummelkamp TR, Bernards R and Agami R: A
system for stable expression of short interfering RNAs in mammalian
cells. Science. 296:550–553. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Gartel AL, Goufman E, Najmabadi F and
Tyner AL: Sp1 and Sp3 activate p21 (WAF1/CIP1) gene transcription
in the Caco-2 colon adenocarcinoma cell line. Oncogene.
19:5182–5188. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Takami Y, Russell MB, Gao C, Mi Z, Guo H,
Mantyh CR and Kuo PC: Sp1 regulates osteopontin expression in SW480
human colon adenocarcinoma cells. Surgery. 142:163–169. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zhu GH, Lenzi M and Schwartz EL: The Sp1
transcription factor contributes to the tumor necrosis
factor-induced expression of the angiogenic factor thymidine
phosphorylase in human colon carcinoma cells. Oncogene.
21:8477–8485. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Diala I, Wagner N, Magdinier F, Shkreli M,
Sirakov M, Bauwens S, Schluth-Bolard C, Simonet T, Renault VM, Ye
J, et al: Telomere protection and TRF2 expression are enhanced by
the canonical Wnt signalling pathway. EMBO Rep. 14:356–363. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Wu S, Ge Y, Li X, Yang Y, Zhou H, Lin K,
Zhang Z and Zhao Y: BRM-SWI/SNF chromatin remodeling complex
enables functional telomeres by promoting co-expression of TRF2 and
TRF1. PLoS Genet. 16:e10087992020. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Dong W, Wang L, Chen X, Sun P and Wu Y:
Upregulation and CpG island hypomethylation of the TRF2 gene in
human gastric cancer. Dig Dis Sci. 55:997–1003. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Luo Z, Feng X, Wang H, Xu W, Zhao Y, Ma W,
Jiang S, Liu D, Huang J and Songyang Z: Mir-23a induces telomere
dysfunction and cellular senescence by inhibiting TRF2 expression.
Aging Cell. 14:391–399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Dinami R, Ercolani C, Petti E, Piazza S,
Ciani Y, Sestito R, Sacconi A, Biagioni F, le Sage C, Agami R, et
al: miR-155 drives telomere fragility in human breast cancer by
targeting TRF1. Cancer Res. 74:4145–4156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Dinami R, Porru M, Amoreo CA, Sperduti I,
Mottolese M, Buglioni S, Marinelli D, Maugeri-Saccà M, Sacconi A,
Blandino G, et al: TRF2 and VEGF-A: An unknown relationship with
prognostic impact on survival of colorectal cancer patients. J Exp
Clin Cancer Res. 39:1112020. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Zizza P, Dinami R, Porru M, Cingolani C,
Salvati E, Rizzo A, D'Angelo C, Petti E, Amoreo CA, Mottolese M, et
al: TRF2 positively regulates SULF2 expression increasing VEGF-A
release and activity in tumor microenvironment. Nucleic Acids Res.
47:3365–3382. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Gavert N and Ben-Ze'ev A: Beta-Catenin
signaling in biological control and cancer. J Cell Biochem.
102:820–828. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Dhillon AS, Hagan S, Rath O and Kolch W:
MAP kinase signalling pathways in cancer. Oncogene. 26:3279–3290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Jones SM and Kazlauskas A: Growth
factor-dependent signaling and cell cycle progression. FEBS Lett.
490:110–116. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Roberts EC, Shapiro PS, Nahreini TS, Pages
G, Pouyssegur J and Ahn NG: Distinct cell cycle timing requirements
for extracellular signal-regulated kinase and phosphoinositide
3-kinase signaling pathways in somatic cell mitosis. Mol Cell Biol.
22:7226–7241. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Nijjar T, Bassett E, Garbe J, Takenaka Y,
Stampfer MR, Gilley D and Yaswen P: Accumulation and altered
localization of telomere-associated protein TRF2 in immortally
transformed and tumor-derived human breast cells. Oncogene.
24:3369–3376. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Picco V, Coste I, Giraud-Panis MJ, Renno
T, Gilson E and Pagès G: ERK1/2/MAPK pathway-dependent regulation
of the telomeric factor TRF2. Oncotarget. 7:46615–46627. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Shukla S, Kanwal R, Shankar E, Datt M,
Chance MR, Fu P, MacLennan GT and Gupta S: Apigenin blocks IKKα
activation and suppresses prostate cancer progression. Oncotarget.
6:31216–31232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Li T, Wong VK, Jiang ZH, Jiang SP, Liu Y,
Wang TY, Yao XJ, Su XH, Yan FG, Liu J, et al: Mutation of cysteine
46 in IKK-beta increases inflammatory responses. Oncotarget.
6:31805–31819. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
An J, Wu M, Xin X, Lin Z, Li X, Zheng Q,
Gui X, Li T, Pu H, Li H and Lu D: Inflammatory related gene IKKα,
IKKβ, IKKγ cooperates to determine liver cancer stem cells
progression by altering telomere via heterochromatin protein
1-HOTAIR axis. Oncotarget. 7:50131–50149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Baskar R, Dai J, Wenlong N, Yeo R and Yeoh
KW: Biological response of cancer cells to radiation treatment.
Front Mol Biosci. 1:242014. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Saha A, Shree Padhi S, Roy S and Banerjee
B: HCT116 colonospheres shows elevated expression of hTERT and
β-catenin protein-a short report. J Stem Cells. 9:243–251.
2014.PubMed/NCBI
|
|
149
|
Saha A, Roy S, Kar M, Roy S, Thakur S,
Padhi S, Akhter Y and Banerjee B: Role of telomeric TRF2 in
orosphere formation and CSC phenotype maintenance through efficient
DNA repair pathway and its correlation with recurrence in OSCC.
Stem Cell Rev Rep. 14:871–887. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Janoušková E, Nečasová I, Pavloušková J,
Zimmermann M, Hluchý M, Marini V, Nováková M and Hofr C: Human Rap1
modulates TRF2 attraction to telomeric DNA. Nucleic Acids Res.
43:2691–2700. 2015. View Article : Google Scholar
|
|
151
|
Anuja K, Chowdhury AR, Saha A, Roy S, Rath
AK, Kar M and Banerjee B: Radiation-induced DNA damage response and
resistance in colorectal cancer stem-like cells. Int J Radiat Biol.
95:667–679. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Osterwald S, Deeg KI, Chung I, Parisotto
D, Wörz S, Rohr K, Erfle H and Rippe K: PML induces compaction,
TRF2 depletion and DNA damage signaling at telomeres and promotes
their alternative lengthening. J Cell Sci. 128:1887–1900. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Zhang P, Pazin MJ, Schwartz CM, Becker KG,
Wersto RP, Dilley CM and Mattson MP: Nontelomeric TRF2-REST
interaction modulates neuronal gene silencing and fate of tumor and
stem cells. Curr Biol. 18:1489–1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Preusser M, de Ribaupierre S, Wöhrer A,
Erridge SC, Hegi M, Weller M and Stupp R: Current concepts and
management of glioblastoma. Ann Neurol. 70:9–21. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Bao S, Wu Q, McLendon RE, Hao Y, Shi Q,
Hjelmeland AB, Dewhirst MW, Bigner DD and Rich JN: Glioma stem
cells promote radioresistance by preferential activation of the DNA
damage response. Nature. 444:756–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Bai Y, Lathia JD, Zhang P, Flavahan W,
Rich JN and Mattson MP: Molecular targeting of TRF2 suppresses the
growth and tumorigenesis of glioblastoma stem cells. Glia.
62:1687–1698. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Simonet T, Zaragosi LE, Philippe C,
Lebrigand K, Schouteden C, Augereau A, Bauwens S, Ye J,
Santagostino M, Giulotto E, et al: The human TTAGGG repeat factors
1 and 2 bind to a subset of interstitial telomeric sequences and
satellite repeats. Cell Res. 21:1028–1038. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Cherfils-Vicini J, Iltis C, Cervera L,
Pisano S, Croce O, Sadouni N, Győrffy B, Collet R, Renault VM,
Rey-Millet M, et al: Cancer cells induce immune escape via
glycocalyx changes controlled by the telomeric protein TRF2. EMBO
J. 38:e1000122019. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Talmadge JE and Gabrilovich DI: History of
myeloid-derived suppressor cells. Nat Rev Cancer. 13:739–752. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Karlseder J, Broccoli D, Dai Y, Hardy S
and de Lange T: p53- and ATM-dependent apoptosis induced by
telomeres lacking TRF2. Science. 283:1321–1325. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Pal D, Sharma U, Singh SK, Kakkar N and
Prasad R: Over-expression of telomere binding factors (TRF1 &
TRF2) in renal cell carcinoma and their inhibition by using SiRNA
induce apoptosis, reduce cell proliferation and migration in vitro.
PLoS One. 10:e01156512015. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Bidzinska J, Baginski M and Skladanowski
A: Novel anticancer strategy aimed at targeting shelterin complexes
by the induction of structural changes in telomeric DNA: Hitting
two birds with one stone. Curr Cancer Drug Targets. 14:201–216.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Horikawa I, Fujita K and Harris CC: p53
governs telomere regulation feedback too, via TRF2. Aging (Albany
NY). 3:26–32. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Yang X, Li Z, Yang L, Lei H, Yu H, Liao Z,
Zhou F, Xie C and Zhou Y: Knockdown of telomeric repeat binding
factor 2 enhances tumor radiosensitivity regardless of telomerase
status. J Cancer Res Clin Oncol. 141:1545–1552. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Cabuy E, Newton C, Joksic G, Woodbine L,
Koller B, Jeggo PA and Slijepcevic P: Accelerated telomere
shortening and telomere abnormalities in radiosensitive cell lines.
Radiat Res. 164:53–62. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
166
|
Zhong YH, Liao ZK, Zhou FX, Xie CH, Xiao
CY, Pan DF, Luo ZG, Liu SQ and Zhou YF: Telomere length inversely
correlates with radiosensitivity in human carcinoma cells with the
same tissue background. Biochem Biophys Res Commun. 367:84–89.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Orun O, Tiber PM and Serakinci N: Partial
knockdown of TRF2 increase radiosensitivity of human mesenchymal
stem cells. Int J Biol Macromol. 90:53–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Ning HB, Li JC, Liu ZG and Fan DM: DNA
damage increases telomerase activity and mRNA expression of
telomeric repeat binding factor 2 in gastric cancer cells.
Available in: www.cnki.net. World Chin J Digest. 14:942–946. 2006.
View Article : Google Scholar
|
|
169
|
Ning HB, Wang YH, Zhang LF, et al:
Reversal of multidrug resistance in gastric cancer cells by
telomeric repeat binding factor 2 small interfering RNA. Available
in: www.cnki.net. Chin J Diges. 31:481–483. 2011.
|
|
170
|
Benhamou Y, Picco V, Raybaud H, Sudaka A,
Chamorey E, Brolih S, Monteverde M, Merlano M, Lo Nigro C,
Ambrosetti D and Pagès G: Telomeric repeat-binding factor 2: A
marker for survival and anti-EGFR efficacy in oral carcinoma.
Oncotarget. 7:44236–44251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Roy S, Roy S, Kar M, Thakur S, Akhter Y,
Kumar A, Delogu F, Padhi S, Saha A and Banerjee B: p38 MAPK pathway
and its interaction with TRF2 in cisplatin induced chemotherapeutic
response in head and neck cancer. Oncogenesis. 7:532018. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Apetoh L, Végran F, Ladoire S and
Ghiringhelli F: Restoration of antitumor immunity through selective
inhibition of myeloid derived suppressor cells by anticancer
therapies. Curr Mol Med. 11:365–372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Bruchard M, Mignot G, Derangère V, Chalmin
F, Chevriaux A, Végran F, Boireau W, Simon B, Ryffel B, Connat JL,
et al: Chemotherapy-triggered cathepsin B release in
myeloid-derived suppressor cells activates the Nlrp3 inflammasome
and promotes tumor growth. Nat Med. 19:57–64. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Sevko A, Michels T, Vrohlings M, Umansky
L, Beckhove P, Kato M, Shurin GV, Shurin MR and Umansky V:
Antitumor effect of paclitaxel is mediated by inhibition of
myeloid-derived suppressor cells and chronic inflammation in the
spontaneous melanoma model. J Immunol. 190:2464–2471. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Wang M, Cao J, Zhu JY, Qiu J, Zhang Y, Shu
B, Ou TM, Tan JH, Gu LQ, Huang ZS, et al: Curcusone C induces
telomeric DNA-damage response in cancer cells through inhibition of
telomeric repeat factor 2. Biochim Biophys Acta Proteins Proteom.
1865:1372–1382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Jiao Y, Zhang W, Liu J, Ni W, Xu W, Jin J
and Qian W: Telomere attrition and chromosome instability via
downregulation of TRF2 contributes to arsenic trioxide-induced
apoptosis of human T-Cell leukemia cell line molt-4 cells. Cancer
Biol Ther. 6:1186–1192. 2007. View Article : Google Scholar : PubMed/NCBI
|














