Introduction
Hypopharyngeal squamous cell carcinoma (HSCC) is a
type of cancer that originates in the cells lining the bottom of
the throat, known as the hypopharynx, which connects the esophagus
to the larynx. Although this type of cancer represents only a small
fraction of all head and neck cancers, its prognosis is markedly
worse compared with other types of head and neck cancer,
characterized by an overall 5-year survival rate of ~30 to 35%
(1). There are three main treatment
options for hypopharyngeal cancer: i) Surgery, ii) radiotherapy and
iii) chemotherapy; however, the optimal therapy regimen depends
mainly on the stage of the cancer. Recently, a multidisciplinary
regimen involving surgery or radiotherapy with chemotherapy
simultaneously has been recommended to treat advanced HSCC
(2). Due to its specific
localization near the larynx, chemoradiotherapy is preferred over
surgical resection with adjuvant chemotherapy, to preserve
laryngeal functions, including breathing, swallowing and speaking
(2). In this context, regardless of
the selected regimen, in HSCC, chemotherapy can be beneficial for
shrinking tumor lesions, mitigating locoregional recurrence and
improving disease-free survival (3), thereby highlighting the need for novel
chemotherapeutic drugs with high efficacy.
Caffeic acid phenethyl ester (CAPE) is the primary
biologically active phenolic compound found in honeybee propolis
and has been documented to have anti-inflammatory, antioxidant,
antiviral and anti-microbial features (4). CAPE has also been identified as a
prospective anticancer drug candidate due to its selective
cytotoxicity toward cancer cells (5). Its therapeutic versatility has been
demonstrated based on its efficacy as a chemotherapeutic adjuvant,
as it potentiates the effectiveness of therapy while mitigating the
chemotherapy-induced side effects in various cancers (6). Moreover, CAPE treatment also inhibits
the proliferation, survival and invasion of oral cancer cells by
inhibiting various signaling pathways, such as EGFR, Akt and NF-κB
signaling (7–9). However, further exploration of the
antitumor effects of CAPE specifically on hypopharyngeal cancer
remains necessary.
Inhibitors of apoptosis proteins (IAPs) comprise a
family of proteins that controls cell death. Survivin and X-linked
IAP (XIAP) are the essential members of the IAP family; they exert
inhibitory effects on caspase-activity and are recognized as
promising therapeutic targets of cancer therapy due to their
overexpression in a variety of cancers (10,11).
Survivin, which is well known for its ability to inhibit apoptosis
and regulate the cell cycle, exhibits high expression in most
cancers and is associated with tumor aggressiveness and unfavorable
clinical outcomes (12).
Furthermore, the expression of survivin has been linked to a poor
prognosis and short overall survival, rendering it a possible
diagnostic marker for head and neck squamous cell carcinoma
(13,14). XIAP is the most powerful member of
the IAP family, as it directly interacts with, and counteracts
caspases 3, 7 and 9 (15). It has
been documented that XIAP expression appears to represent a
negative predictor of prognosis in head and neck cancer (16) and in laryngeal carcinoma (17). Moreover, survivin and XIAP can be
combined to form a complex known as IAP-IAP, in which they work
together to counteract the activity of caspase-9 (18). Thus, it may be a promising approach
to develop novel medications that concurrently target survivin and
XIAP during cancer therapy.
In the present study, the pharmacological activity
of CAPE regarding the induction of intrinsic mitochondrial
apoptosis, which was ascribed to the concurrent inhibition of the
survivin and XIAP proteins was assessed, suggesting that CAPE is a
potent drug candidate in the therapy of HSCC.
Materials and methods
Cell culture and reagents
The FaDu and SNU-1041 human HSCC cell lines were
purchased from the Korean Cell Line Bank (Seoul, Republic of Korea)
and cultured in either Minimum Essential Medium (FaDu) or RPMI-1640
medium (SNU-1041) from Welgene, Inc. containing 10% FBS (Welgene,
Inc.) and 1% penicillin/streptomycin (P/S). The IHOK cell line was
kindly provided by Yonsei University (Seoul, Republic of Korea) and
cultured in KBM™ Gold Keratinocyte Growth Basal Medium (Lonza
Group, Ltd.) supplemented with KGMTM-2 Keratinocyte Growth Medium-2
SingleQuots™ Supplements and Growth Factors (Lonza Group, Ltd.).
The cells were incubated at 37°C in a humidified environment with
5% CO2. All experiments were conducted when the cells
had grown to ~50% confluence. All chemicals were dissolved in DMSO
and stored at −20°C. CAPE was purchased from Santa Cruz
Biotechnology, Inc. Each cell line was divided into two groups for
further analysis of cellular responses to CAPE treatment: i) The
vehicle control group and ii) the CAPE (70 µM for FaDu and 100 µM
for SNU-1041 and IHOK) treatment group. Vehicle- and CAPE-treated
cells were incubated at 37°C for 24 h. For pre-treatment of
inhibitors for 1 h (Z-VAD-FMK) or 2 h (CHX and MG132) at 37°C, each
cell line was divided into following two groups: i) Vehicle control
group and ii) Z-VAD-FMK (10 µM), CHX (100 ng/ml for FaDu and 50
ng/ml for SNU-1041), and MG132 (800 nM) treatment group.
Trypan blue assay
FaDu, SNU-1041 and IHOK cells at 50% confluency were
treated with vehicle or CAPE. Trypan blue solution in a
concentration of 0.4% (Corning, Inc.) was used to stain the cells
at a 1:1 ratio at room temperature (RT). Immediately, after
staining, cell viability was evaluated automatically using a
CytoSMART cell counter (Corning, Inc.).
Soft agar assay
Basal Medium Eagle (BME; Sigma-Aldrich,) was
dissolved in distilled water (DW) supplemented with sodium
bicarbonate, resulting in a 2X BME solution, then filtered using a
0.2-µm filter (Sartorius AG). A mixture containing 1.25% agar, 2X
BME, FBS, PBS, l-glutamine and gentamicin was then prepared. A
total of 3 ml of the 1.25% agar mixture containing the indicated
amounts of CAPE (70 µM for FaDu and 100 µM for SNU-1041) was added
to each well of the 6-well plates. The agar mixture was then
allowed to solidify for 2 h at RT. Subsequently, a 10% BME solution
was prepared by diluting 2X BME with DW and supplementing with
l-glutamine, gentamicin and FBS. FaDu and SNU-1041 cells were
suspended in the 10% BME solution and mixed with 1.25% agar. After
mixing, 1 ml of the mixture containing 0.8×104 cells was
added directly onto the 6-well solid-bottom agar plates. The plates
were incubated at RT for 2 h for agar solidification and then
placed in a humidified incubator at 37°C with 5% CO2 for
a period of ~4 weeks. The 6-well plates were supplemented once a
week with 150 µl of complete medium containing the indicated
concentrations of CAPE, to prevent agar desiccation. Images of cell
colonies were captured using a CKX53 microscope (Olympus
Corporation) and colony counts were analyzed automatically using
the ImageJ software, version 1.53t (National Institutes of
Health).
Sphere formation assay
Cells were suspended in a serum-free medium
supplemented with 0.01X N-2 and B-27 supplements, 25 ng/ml of human
basic fibroblast growth factor (bFGF; Invitrogen; Thermo Fisher
Scientific, Inc.) and human epidermal growth factor (EGF; Thermo
Fisher Scientific, Inc.) and 1% P/S. The cells (1×104
cells/well) were cultured in ultralow attachment 6-well plates
(Corning, Inc.) for 5 days (FaDu) and 11 days (SNU-1041). Spheres
were randomly photographed using an inverted light microscope
(Nikon Corporation) and counted automatically using ImageJ
software, version 1.53t.
Evaluation of nuclear morphological
changes
FaDu and SNU-1041 cells, which were treated with
either a vehicle or CAPE when they reached ~50% confluence were
fixed with 70% ethanol at −20°C overnight and stained with a DAPI
solution (2 µg/ml; MilliporeSigma) on a glass slide for 10 min at
RT. Images of the nuclear morphological changes occurring in
apoptotic cells were captured by fluorescence microscopy (Leica
DMi8; Leica Microsystems GmbH).
Measurement of cell-cycle
distribution
After treating the cells with vehicle or CAPE when
they reached ~50% confluence, FaDu and SNU-1041 cells were fixed
with 70% ethanol overnight at −20°C and stained with a propidium
iodide (PI) solution (20 µg/ml; MilliporeSigma) containing RNase A
(20 µg/ml; Thermo Fisher Scientific Inc.) at 37°C for 15 min.
Cell-cycle distribution was then assessed by an LSRFortessa X-20
flow cytometer (BD Biosciences) and analyzed using the FlowJo
software, version 10.8.1 (FlowJo LLC).
Determination of apoptotic cell
populations
FaDu and SNU-1041 cells, which were treated with
either a vehicle or CAPE when they reached approximately 50%
confluence were stained with FITC Annexin V Apoptosis Detection Kit
(cat. no. 556547; BD Pharmingen; BD Biosciences), according to
manufacturer's instructions. In brief, the cells were stained with
Annexin V for 15 min at RT and then stained with PI on ice just
before evaluation using a flow cytometer. The apoptotic population
was evaluated using an LSRFortessa X-20 flow cytometer (BD
Biosciences) and analyzed using the FlowJo software, version
10.8.1.
Western blot analysis
Total protein lysates were extracted using 1X RIPA
buffer (MilliporeSigma) including cOmplete™, Mini protease
inhibitor cocktails (Roche Diagnostics) and Pierce™ phosphatase
inhibitor tablets (Thermo Scientific; Thermo Fisher Scientific,
Inc.). A quantitative analysis of protein concentrations was
carried out using a DC Protein Assay Kit (Bio-Rad Laboratories,
Inc.). Equal amounts of protein were loaded in each lane of the
gel, but this amount was adjusted according to the protein
examined. Protein lysates were separated using SDS-PAGE. The
separated proteins were then electrotransferred onto a PVDF
membrane. The membrane was blocked with 5% skim milk in TBST
including 0.05% Tween 20 (Duchefa Biochemie) for 1 h 30 min at RT,
to prevent non-specific binding of antibodies to the membrane.
Subsequently, the membrane was thoroughly rinsed with TBST,
incubated with the designated primary antibodies overnight at 4°C,
then exposed to HRP-conjugated secondary antibodies for 4 h at 4°C.
The protein bands of interest were visualized using the WestGlow™
PICO PLUS chemiluminescent Substrate (BIOMAX) on an X-ray film or
using the Image Quant LAS 500 system (Cytiva). β-actin was used as
a loading control. The intensity of protein bands was quantified
using ImageJ software, version 1.53t. Information concerning the
antibodies used as well as the amount of loaded protein and the
percentages of the gels for the experiments conducted are presented
in Table I.
 | Table I.List of antibodies and conditions
used for western blot analysis. |
Table I.
List of antibodies and conditions
used for western blot analysis.
| Antibody name | Source | Company (cat.
no.) | Dilution | Gel percentage
(%) | Protein loading
amount |
|---|
| c-caspase-3 | Rabbit | Cell Signaling
Technology, Inc. (#9664) | 1:1,000 | 15 | 25 µg |
| c-PARP | Rabbit | Cell Signaling
Technology, Inc. (#9541) | 1:2,000 | 8 | 25 µg |
| c-caspase-9 | Rabbit | Cell Signaling
Technology, Inc. (#9501) | 1:2,000 | 12 | 30 µg |
| c-caspase-8 | Rabbit | Cell Signaling
Technology, Inc. (#9496) | 1:2,000 | 12 | 40 µg |
| Bid (tBid) | Rabbit | Cell Signaling
Technology, Inc. (#2002) | 1:1,000 | 15 | 25 µg |
| Cytochrome
c | Mouse | BD Pharmingen
(#556433) | 1:1,000 | 15 | 30 µg |
| Cox IV | Rabbit | Abcam
(#ab16056) | 1:1,000 | 15 | 15 µg |
| α-Tubulin | Mouse | Santa Cruz
Biotechnology, Inc. (#sc-5286) | 1:3,000 | 12 | 15 µg |
| Survivin | Mouse | Cell Signaling
Technology, Inc. (#2802) | 1:1,000 | 15 | 25 µg |
| XIAP | Rabbit | Cell Signaling
Technology, Inc. (#14334) | 1:3,000 | 12 | 20 µg |
| β-actin | Mouse | Santa Cruz
Biotechnology, Inc. (#sc-47778) | 1:3,000 | 12 | 15 µg |
| Goat Anti-Rabbit
IgG antibody (HRP) |
| GeneTex, Inc.
(#GTX213110-01) | 1:3,000 |
|
|
| Goat Anti-Mouse IgG
antibody (HRP) |
| GeneTex, Inc.
(#GTX213111-01) | 1:3,000 |
|
|
Assessment of changes on the
mitochondrial membrane potential (ΔΨm)
The effects of CAPE on the ΔΨm were determined using
a MitoScreen JC-1 kit (cat. no. 551302; BD Pharmingen; BD
Biosciences), according to the manufacturer's protocols. Briefly,
FaDu and SNU-1041 cells were washed twice with PBS and incubated
with a 1X JC-1 working solution for 30 min at 37°C. The stained
cells were washed and resuspended in 1X assay buffer, then analyzed
using an LSRFortessa X-20 flow cytometer (BD Biosciences) and
interpreted using the FlowJo software, version 10.8.1.
Cytosolic and mitochondrial
fractionation
Vehicle- or CAPE-treated FaDu and SNU-1041 cells
were separated into cytosolic and mitochondrial extracts using the
Mitochondria/Cytosol Fractionation Kit (cat. no. ab65320; Abcam),
according to the manufacturer's protocols. In brief, the cells were
resuspended in 1X cytosol extraction buffer mix including protease
inhibitors and DTT, vortexed for 2 min and placed on ice for 10
min, to allow complete disruption of cell membranes. Following
centrifugation for 15 min at −10,000 × g at 4°C, the supernatants
(including the cytosolic fraction of the cells) were carefully
transferred to new microtubes. The remaining cell pellets were
washed with PBS, resuspended in 1X mitochondrial extraction buffer
mix containing protease inhibitors and DTT, vortexed for 10 sec,
and centrifuged using the aforementioned conditions. The resulting
supernatants (including the mitochondrial fraction of the cells)
were collected in new microtubes.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® reagent (Thermo Fisher
Scientific, Inc.) was utilized to extract total RNA from FaDu and
SNU-1041 cells and 1 µg of the extracted RNA was reverse
transcribed into cDNA using the AMPIGENE cDNA Synthesis Kit (Enzo
Life Sciences, Inc.): 42°C for 30 min and 85°C for 10 min. The
resulting cDNA was used as the input for PCR using the AMPIGENE
qPCR Green Mix Hi-Rox reagent (Enzo Life Sciences, Inc.). Real-time
PCR was carried out on an Applied Biosystems StepOne Plus Real-Time
PCR System (Applied Biosystems) using the following conditions for
all genes: 95°C for 2 min, followed by 40 cycles at 95°C for 10 sec
and 60°C for 30 sec. The amount of the GAPDH gene was used
to normalize the relative amount of each gene, which was measured
using the 2−ΔΔCq method (19). The qPCR primers used in this
experiment were as follows: Survivin forward,
5′-ACTTGGCCCAGTGTTTCTT-3′ and reverse, 5′-GACAGAAAGGAAAGCGCAAC-3′;
XIAP forward, 5′-TGGTATCCAGGGTGCAAATATC-3′ and reverse,
5′-GTTCTTACCAGACACTCCTCAAG-3′; and GAPDH forward,
5′-GTGGTCTCCTCTGACTTCAAC-3′ and reverse,
5′-CCTGTTGCTGTAGCCAAATTC-3′. The primers were designed based on the
NCBI-BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) reference
sequences: Survivin-CR541740.1, XIAP-BC032729.1 and
GAPDH-BC083511.1. The detailed primer binding sites of respective
proteins are included in the Fig.
S1, Fig. S2, Fig. S3.
Statistical analysis
All graphs were generated using GraphPad Prism,
version 8.0 (GraphPad Software; Dotmatics) and all statistical
analyses were performed using SPSS, version 26.0 (IBM, Corp). The
results of three independent biological experiments are presented
as the average (mean) value together with the measure of variation
[standard deviation (SD)] for all data. Statistical significance
was determined by conducting either an unpaired two-tailed
Student's t-test or a one-way ANOVA with Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
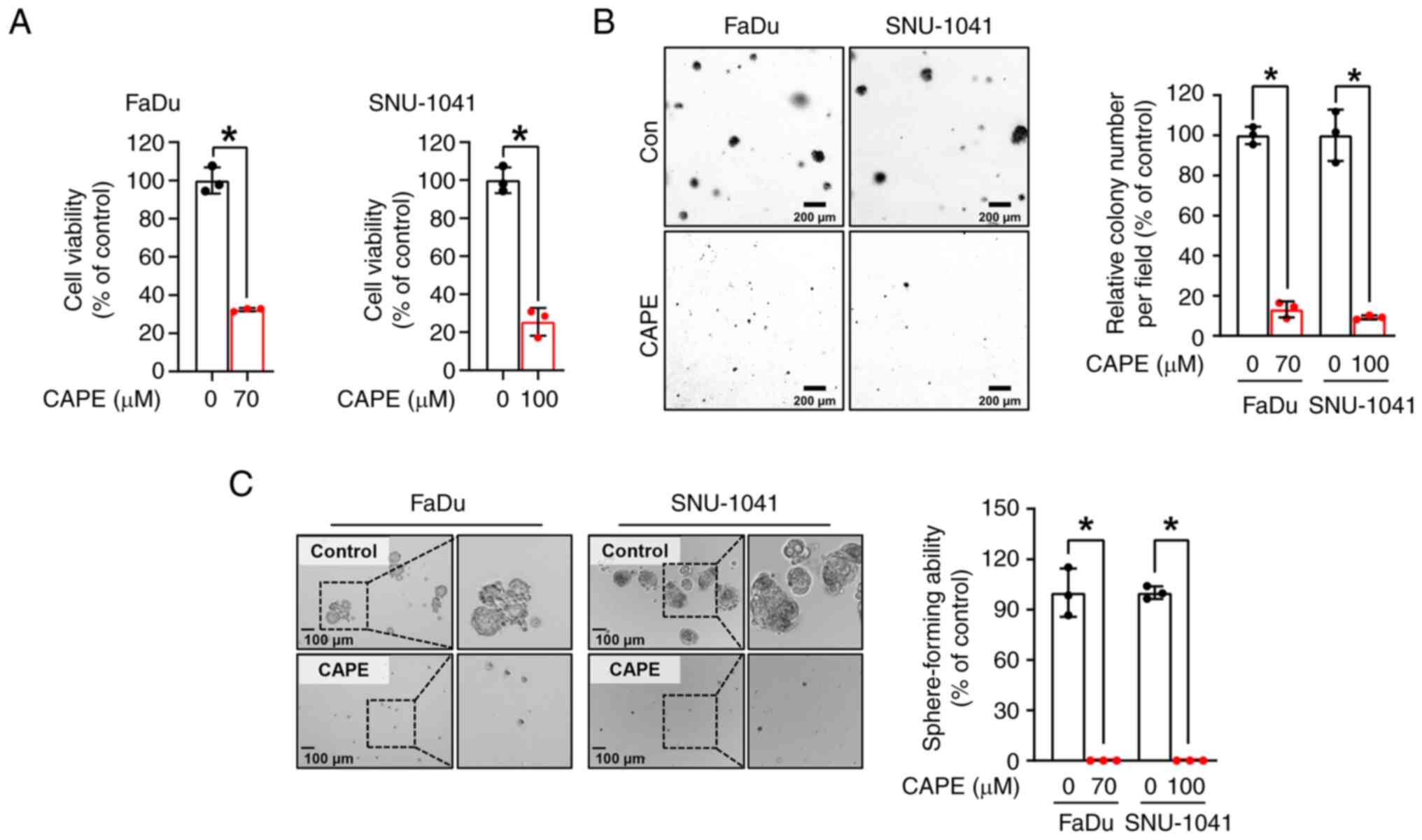
CAPE inhibits the anchorage-dependent
and independent growth of HSCC cell lines
To examine the impacts of CAPE on HSCC cell growth,
a trypan blue assay was performed on two HSCC cell lines, FaDu and
SNU-1041, which had been treated with CAPE for 24 h. As revealed in
Fig. 1A, the growth of the two cell
lines was significantly reduced after treatment with CAPE, but not
with the vehicle control. Notably, it was observed that CAPE worked
less effectively in the IHOK cell line, immortalized human oral
keratinocytes, compared with HSCC cell lines, suggesting that CAPE
could selectively attenuate the growth of cancer cells rather than
affect normal cells (Fig. S4). A
soft agar assay was then conducted to further evaluate the
influence of CAPE on the anchorage-independent growth of the HSCC
cell lines. As displayed in Fig.
1B, the clonogenic activity of the two cell lines was
attenuated by CAPE treatment, as indicated by the reduced size and
number of the cell colonies. Furthermore, to examine the tumor
suppressive capability of CAPE in a 3D culture system, a
sphere-formation assay was carried out. As demonstrated in Fig. 1C, the sphere-forming ability of both
cell lines was strongly inhibited in CAPE-treated cells compared
with control cells. These findings suggested that CAPE thwarts cell
growth and carcinogenesis in these HSCC cell lines.
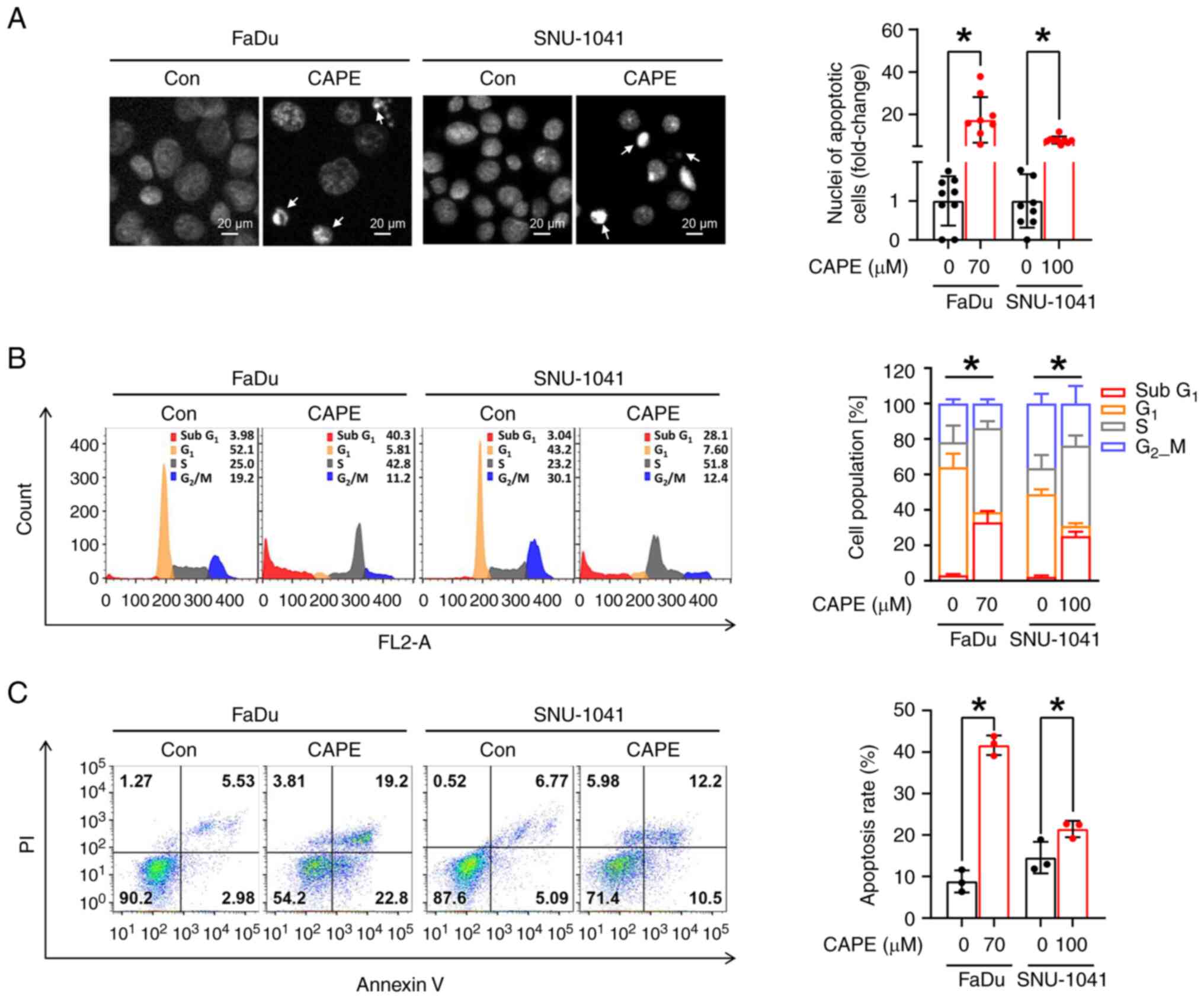
CAPE impedes tumorigenicity by
triggering caspase-dependent apoptosis in HSCC cell lines
To confirm whether the observed cytotoxic effect of
CAPE on HSCC cell lines was caused by the activation of apoptotic
signaling, three apoptosis detection assays were employed. First,
the morphological changes in the nuclei of CAPE-treated cells
compared with vehicle-treated cells were evaluated; fragmented or
condensed nuclei were observed, which are regarded as typical
morphological signs of apoptosis (Fig.
2A). Subsequently, whether cell populations within the
sub-G1 phases expanded after CAPE treatment was
investigated. As illustrated in Fig.
2B, a significant increase in the sub-G1 population
in both cell lines was observed after the application of CAPE.
Finally, apoptotic cell populations were measured using annexin/PI
double staining, to investigate further the effect of CAPE on the
induction of apoptosis. It was also revealed that CAPE treatment
enhanced the populations within the early and late apoptotic
compartments (Fig. 2C). Based on
these results, whether the apoptosis stimulated in the cells
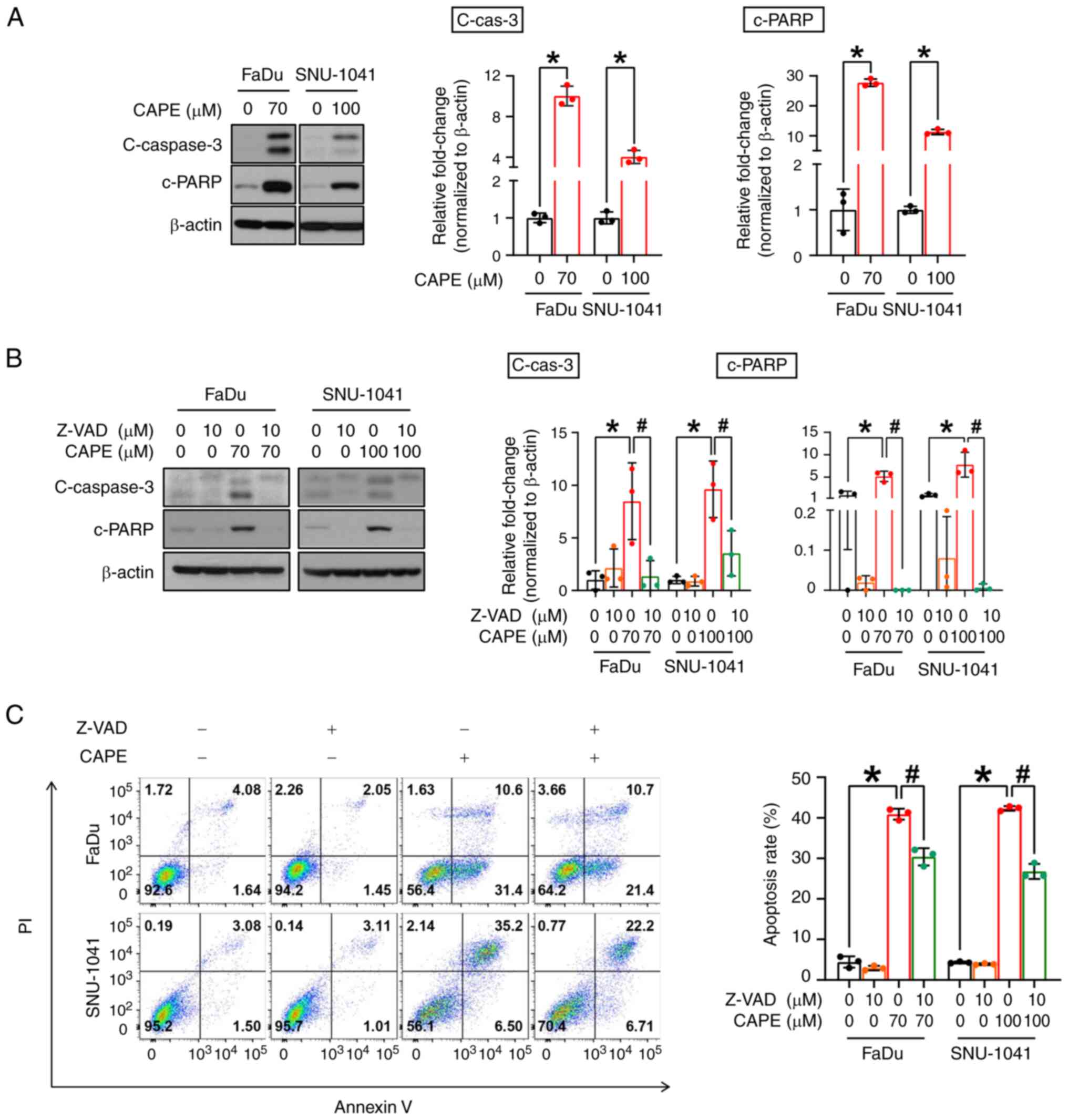
exposed to CAPE was dependent on the activation of caspase
signaling, was examined. As shown in Fig. 3A, the CAPE treatment notably
stimulated the formation of cleaved forms of caspase-3
(c-caspase-3) and induced the expression of cleaved
poly(ADP-ribose) polymerase (c-PARP). To obtain evidence to
corroborate these results, the cells were treated with Z-VAD-FMK,
which is a caspase-inhibitor, for 1 h before exposing them to CAPE
treatment for 24 h, to investigate whether CAPE can induce
caspase-dependent apoptosis in the two cell lines. As expected, the
induction of the expression of c-caspase-3 and c-PARP and annexin
V-positive compartments by the CAPE treatment was reversed by
pre-treatment of the cells with Z-VAD-FMK (Fig. 3B and C). Taken together, these
results indicated that CAPE considerably attenuates the growth of
HSCC cells, possibly by triggering caspase-dependent apoptotic
processes.
CAPE exerts cell-killing effects
through the activation of mitochondrial apoptotic signaling in HSCC
cell lines
Apoptosis can be triggered via two primary pathways,
the extrinsic or intrinsic pathway, with formation of cleaved
caspase-8 and caspase-9 being considered a typical marker of each
pathway, respectively. Therefore, the pathway which was responsible
for the apoptotic cell-killing effect in cells that were subjected
to CAPE treatment was investigated. After exposure of both cell
lines to CAPE for 24 h, an increase in the expression of cleaved
caspase-9 (c-caspase-9), but not of cleaved caspase-8
(c-caspase-8), Bid, and t-Bid were observed, indicating that CAPE
may promote intrinsic apoptosis signaling (Fig. 4A). In the intrinsic pathway,
mitochondria lose their ΔΨm and exhibit increased membrane
permeabilization, which causes the release of proteins from the
mitochondrial intermembrane spaces, such as cytochrome c
(20). To demonstrate further the
impact of CAPE on the activation of the intrinsic apoptotic
pathway, changes in ΔΨm were measured by employing the JC-1
fluorescent dye. As expected, the number of JC-1 aggregates (red
fluorescence) was decreased in cells treated with CAPE, indicating
that the treatment caused a reduction in the ΔΨm (Fig. 4B). Next, it was investigated whether
the changes in the ΔΨm caused by CAPE led to the outer
mitochondrial membrane permeabilization, causing the release of
cytochrome c. Cytochrome c oxidase subunit 4I1 (Cox
IV), a cytochrome c oxidase enzyme located within the inner
membrane of the mitochondria (21),
and α-tubulin, a cytoplasmic marker protein (22), were used as mitochondrial and
cytosol loading controls, respectively. As demonstrated in Fig. 4C, it was found that cytochrome
c was transferred from the mitochondria to the cytosol, as
indicated by the decrease in cytochrome c detected in the
mitochondria and the increase in cytochrome c observed in
the cytosol in response to CAPE. Accordingly, the apoptosis
observed in the two CAPE-treated cell lines was attributed to an
intrinsic mitochondrial signaling process.
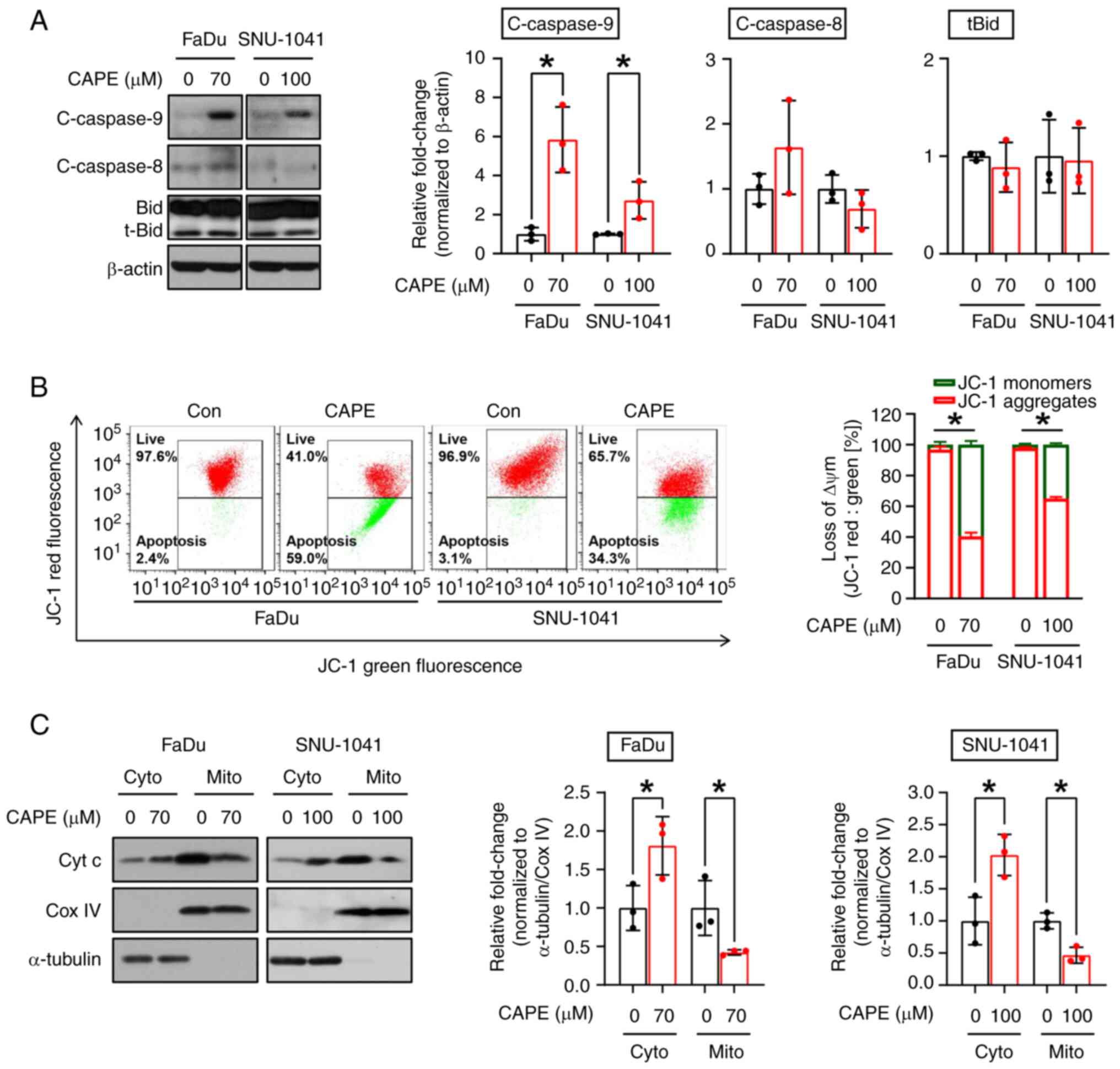 | Figure 4.Effect of CAPE on the mitochondrial
apoptotic pathway in human hypopharyngeal squamous cell carcinoma
cell lines. FaDu and SNU-1041 cells were exposed to DMSO or CAPE
for 24 h. (A) Western blot images showing the expression levels of
c-caspase-9 and c-caspase-8. β-actin was used as an internal
control. (B) JC-1 aggregates in DMSO- or CAPE-treated cells were
measured using fluorescence-activated cell sorting cytometry. (C)
Mitochondrial and cytosolic fractions were prepared to detect
cytochrome c release from mitochondria into the cytosol. Cox
IV and α-tubulin were used as loading controls and as markers of
mitochondria and the cytosol, respectively. All graphs represent
the mean ± SD of three independent experiments. *P<0.05;
unpaired two-tailed Student's t-test. CAPE, caffeic acid phenethyl
ester; cleaved caspase-9, c-caspase-9; cleaved caspase-8,
c-caspase-8; Cox IV, cytochrome c oxidase subunit 4I1; Bid,
BH3 interacting domain death agonist; Con, control; ΔΨm,
mitochondrial membrane potential; Cyt c, cytochrome complex; Cyto,
cytoplasm; Mito, mitochondria. |
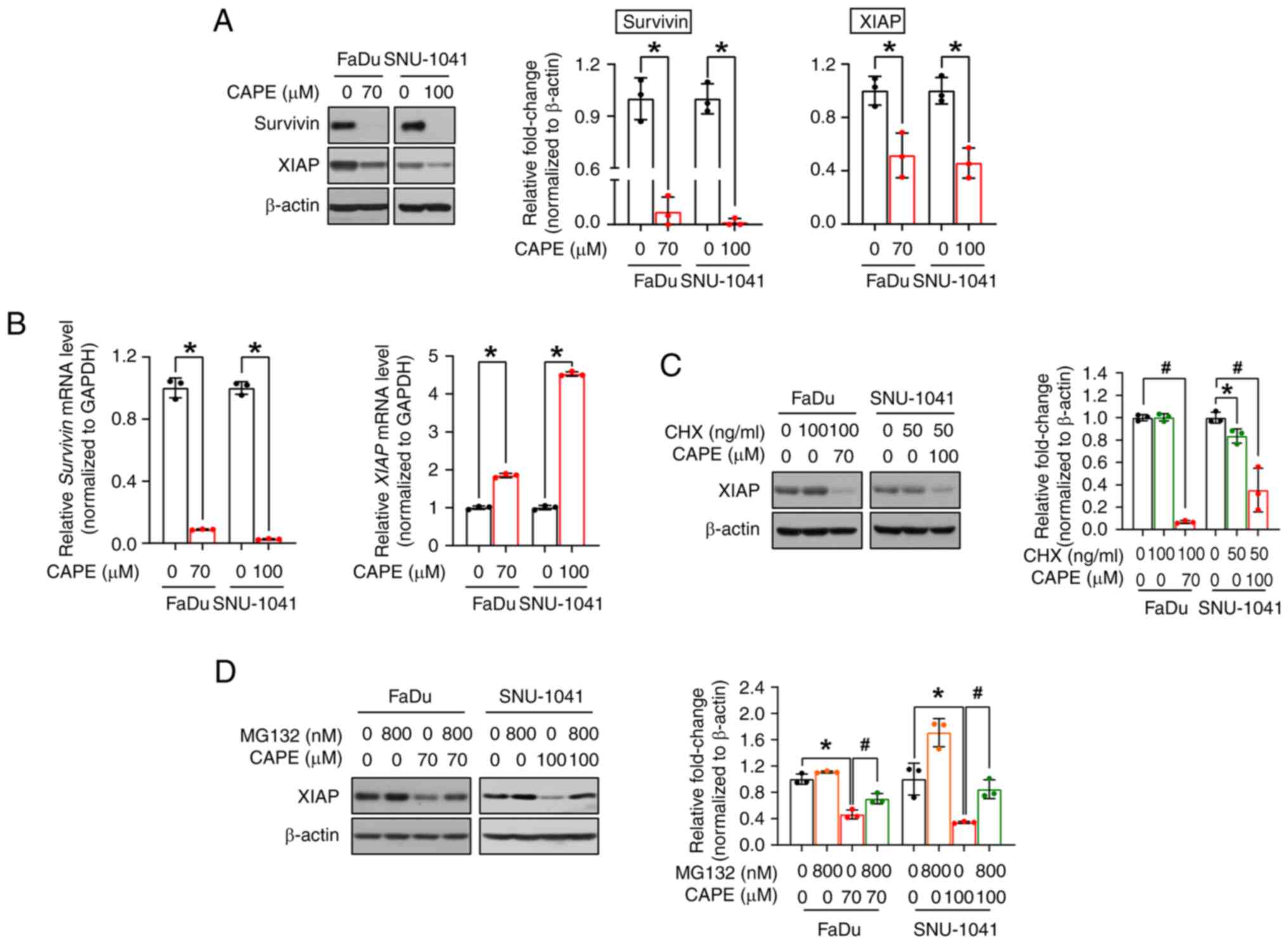
CAPE induces an apoptotic signaling
cascade by blocking the anti-apoptotic proteins survivin and
XIAP
As aforementioned, survivin and XIAP are the major
members of the IAP family, and both hinder apoptosis, leading to
the increased survival of cancer cells (23). Therefore, the expression of survivin
and XIAP were investigated to determine the exact mechanism
underlying CAPE-mediated apoptosis. As expected, CAPE
administration resulted in the downregulation of both proteins
(Fig. 5A). Next, to investigate
whether the changes observed for the two proteins were caused by
transcriptional regulation, the corresponding mRNA expression
levels were measured. As revealed in Fig. 5B, CAPE downregulated the survivin
mRNA, but not the XIAP mRNA, suggesting that survivin alone was
regulated at the transcriptional level. Therefore, it was
hypothesized that XIAP is regulated at the post-translational
level, and it was investigated whether CAPE compromises the
stability of this protein. In advance of implementing cycloheximide
(CHX) and MG132 experiments, a time-course experiment was conducted
to verify the exact time point of XIAP degradation by CAPE and to
minimize its potential effect on cell viability. In Fig. S5, XIAP was downregulated after 6 h
of CAPE treatment in both cell lines. The time was determined when
a 1-h pretreatment of inhibitors was followed by subsequent CAPE
treatment for 6 h. As depicted in Fig.
5C, the level of the XIAP protein was decreased in the two cell
lines after co-treatment with CHX (a protein synthesis inhibitor)
and CAPE, which may indicate that XIAP was controlled by
post-translational regulation, particularly via protein
degradation. Thus, the cells were subsequently treated with MG132
(a proteasome inhibitor) for 2 h prior to CAPE treatment, to
explore whether XIAP protein degradation was accelerated by CAPE in
a proteasome-dependent manner. In Fig.
5D, MG132 treatment restored the downregulated levels of
expression of the XIAP protein in the CAPE-treated groups in both
cell lines. Consequently, these data indicated that CAPE exerts its
apoptosis-inducing effects on HSCC cells by downregulating two
anti-apoptotic proteins, survivin and XIAP, via either
transcriptional or post-translational regulation.
Discussion
In the present study, the potential of the bioactive
component CAPE to treat HSCC via the induction of mitochondrial
apoptosis was demonstrated and was achieved by blocking two potent
anti-apoptotic proteins, survivin and XIAP.
In the present study, cells were initially treated
with both 100 and 120 µM of CAPE for 24 h and it was observed that
120 µM of CAPE significantly decreased the expression of β-actin
indicating excessive cytotoxicity (Fig. S6A). Consequently, the
concentrations were adjusted to a range of 60–100 µM, ensuring that
the expression of β-actin remained constant while c-PARP increased
suitably (Fig. S6B). While a
higher CAPE dose could induce more prominent apoptotic effects, it
also carried a risk of causing excessive cytotoxicity in both
normal and cancer cells. Based on the aforementioned data, 70 µM
CAPE for FaDu and 100 µM CAPE for SNU-1041 were employed, to
demonstrate its anticancer therapeutic potential, while avoiding
any significant effect on the expression of β-actin. Several
previous studies have employed varying concentrations of the same
drug for different cell lines in vitro (24–27).
In the case of CAPE, CAPE was administered at concentrations of
10–50 µM to multiple myeloma cell lines to investigate its
antimyeloma potential (28). A
different study exposed human colorectal cell lines to 75 µM CAPE,
revealing CAPE-induced apoptotic cell death through the inhibition
of survivin (29). Furthermore,
ovarian cancer OV7 cells were treated with 5–100 µM of CAPE to
explore its therapeutic benefits in serous ovarian cancer (30). In a separate study, endometrioid
ovarian carcinoma cells were exposed to CAPE at concentrations
ranging from 1–10 µM, demonstrating its anti-tumorigenic activity
(31). It was also revealed that
CAPE at concentrations ranging from 1–50 µM could induce apoptosis
and oxidative stress in human multiple myeloma cells (32). These findings indicated that in
vitro treatment concentration of some substances may be
dependent on cell context even though they exhibited similar
functions such as apoptosis induction.
Targeting apoptosis has been deemed a successful
approach in cancer therapy as apoptosis evasion is the hallmark of
various cancers, regardless of their etiology and type (33). Apoptosis can be triggered by two
primary pathways such as the binding of death ligands to their
cognate death receptors, which is termed the extrinsic pathway and
cytotoxicity from mitochondrial signaling, which is termed the
intrinsic pathway (34). The
extrinsic pathway relies on Fas, tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) or TNFα to transmit extracellular
signals into cells, which ultimately leads to the activation of
caspase-8. By contrast, in the intrinsic pathway, intercellular
stimuli such as cellular stress, DNA damage and growth factor
withdrawal provoke mitochondrial malfunctioning via the induction
of mitochondrial outer membrane permeabilization, consequently
causing the release of mitochondrial proteins, such as cytochrome
c, SMA and OMI into the cytosol. Caspase-9 and apoptotic
peptidase activating factor 1 (APAF1) are recruited by cytochrome
c to produce cleaved caspase-9, which is the activated form
of caspase-9 (35). These two
apoptotic pathways eventually converge toward the formation of
cleaved caspase-3 and, finally, cleaved PARP, followed by cell
demise (20). Several natural
products exert tumor-inhibitory activities by stimulating the
intrinsic and extrinsic apoptotic signaling pathways exclusively,
respectively, or by triggering the two pathways simultaneously
(36–38). In the latter cellular context, the
intrinsic and extrinsic apoptotic pathways can be integrated
through the truncation of the BH3 interacting domain death agonist
(Bid) protein, which is a process that is facilitated by
c-caspase-8 (39). In the present
study, it was ascertained whether CAPE exclusively triggers one
type of apoptosis or whether it simultaneously induces both types
of apoptosis. It was revealed that the expression of c-caspase-9
alone was increased, whereas the expression of c-caspase-8 and
truncated Bid remained unchanged (Fig.
4A). Based on these results, the authors suggest that CAPE
exerts its HSCC cell-destroying effects mainly via
mitochondria-mediated intrinsic apoptosis.
Since apoptosis is an ongoing and meticulously
controlled mechanism that sustains the cellular equilibrium within
the body of a healthy animal (40),
the expression of c-PARP can be detected at low levels. In Fig. 3A and B, the relative expression
levels of c-PARP in vehicle- and CAPE-treated cells were measured,
and basal protein expression levels were visualized due to an
extended exposure time. However, it is evident that c-PARP
expression levels were further induced by CAPE treatment compared
to basal levels.
Survivin and XIAP are members of the IAP family that
have the potential to regulate cell death, and they have been
identified as reasonable targets for numerous anticancer
therapeutics in various types of cancer (41–44).
Although targeting these proteins individually has been shown to be
an effective intervention per se, a simultaneous therapeutic
intervention against the two proteins appears to be a more feasible
pharmacological strategy (45–48).
As aforementioned, survivin and XIAP may form a survivin-XIAP
complex with a vigorous apoptosis inhibitory activity (18). Thus, targeting both survivin and
XIAP, as CAPE did in the experiments of the present study, may
provide improved treatment options for multiple types of cancers.
In the present study, a CAPE-mediated decrease in the levels of the
survivin protein was observed and appeared to be controlled by
transcriptional regulation, as evidenced by the reduction of the
levels of the survivin mRNA. By contrast, the mRNA level of XIAP
was unexpectedly increased, whereas its protein level was
significantly decreased, by CAPE treatment. Under particular
cellular conditions, the expression of a given protein does not
coincide perfectly with that of its cognate mRNA, possibly due to
the compensatory responses of gene expression, in which the
downregulation of protein levels leads to an upregulation of mRNA
levels, or post-translational modification. In a recent study, it
was observed that the small molecule, bufalin, induced a reduction
in the expression of the E2F2 protein, whereas the expression of
the E2F2 mRNA was increased, suggesting that the downregulation of
the E2F2 protein was a result of post-translational modification
(49). In the present study, it was
also found that CAPE regulated XIAP via a post-translational
modification. Survivin and XIAP complexes enhance the stability of
XIAP by preventing the ubiquitination-mediated degradation of XIAP,
thus resulting in the inhibition of caspase-activity followed by
abnormal tumor growth (50). The
binding of survivin to XIAP can also serve as a salvage mechanism
by blocking the formation of the XIAP-XAF1 complex, which targets
survivin for degradation through the ubiquitin-proteasome system
(51). Because XIAP protein
stability is partly associated with its binding to survivin, the
levels of transcriptionally downregulated survivin cannot prevent
the degradation of XIAP by the proteasome.
Since survivin and XIAP are regulated by different
mechanisms, there may be separate upstream regulators controlling
their expression, respectively. Regarding survivin, the expression
decreased at the mRNA level, suggesting that transcription factors
are likely associated with its downregulation. Previous studies
have reported that the p53 protein could recruit itself alone or in
conjunction with other proteins to the survivin promoter to
suppress it (52,53). Thus, CAPE treatment possibly
mediated the recruitment of p53 to the survivin promoter,
ultimately reducing its expression. On the other hand, XIAP
regulation appears to be executed through post-translational
modifications, particularly proteasome-mediated degradation.
Notably, XIAP can be phosphorylated by Akt and protein kinase C
(PKC), which prevents its ubiquitination and subsequent proteasomal
degradation (54,55). XIAP is also well-known for its E3
ubiquitin ligase activity, allowing it to induce
autoubiquitination. Consequently, it is possible that the
attenuated activation of Akt and PKCs, which cannot shield XIAP
from ubiquitination, or the promotion of XIAP autoubiquitination by
CAPE, could compromise its stability. Therefore, further
investigation of the precise mechanisms underlying the regulation
of survivin and XIAP during CAPE treatment in HSCC remains
necessary.
In previous studies, in vivo experiments
using CAPE have been conducted to evaluate its anticancer activity,
and the results revealed that CAPE has anticancer effects in
hepatocellular carcinoma and colon cancer (56,57).
These findings led to the investigation of whether CAPE may also
inhibit tumor growth in human HSCC in vivo. However, in the
present study, the 3D sphere formation assay was implemented to
demonstrate the anticancer properties of CAPE in cancer stem cell
(CSC) characteristics of HSCC and to explore its potential as a
therapeutic agent, as an alternative to in vivo experiments.
Considering the well-recognized ability of the 3D sphere formation
culture to more effectively recapitulate in vivo tumor
biology compared with the 2D monolayer culture system (58), the data obtained from the 3D culture
system can provide some insights that complement mouse xenograft
experiments. Nevertheless, further investigations are still
warranted to clarify the tumor-inhibitory effects of CAPE on HSCC
in an in vivo setting.
In the clinical field, the preferred initial
treatment approach, as recommended by both American and European
guidelines, typically involves preserving the organ through a
combination of chemotherapy and radiation therapy (59). Given this context, chemotherapy
signifies much more than merely a choice for treating HSCC. The
authors of the present study maintain that CAPE alone may not
eliminate tumors; however, it could serve as a promising
alternative to current therapeutic drugs. This, in turn, could
minimize tumor regions and suppress recurrence, serving as both
neoadjuvant and adjuvant chemotherapy.
Previous studies have explored the anticancer
effects of CAPE on oral cancer, primarily utilizing oral squamous
cell carcinoma cell lines (9,60,61).
In the present study, head and neck cancer cell lines were
employed, specifically FaDu and SNU-1041, which are hypopharyngeal
cancer cell lines technically categorized under nasopharyngeal
cancer. Although oral cancer and hypopharyngeal cancer fall within
the broader classification of head and neck cancer, they possess
distinct staging assessments and treatment standards due to their
unique characteristics and locations within the head and neck
region (62). To the best of the
authors' knowledge, the present study marked the first
demonstration of the cancer-inhibitory efficacy of CAPE through the
induction of cell death in HSCC lines, adding novelty to the study.
However, the authors used only two HSCC cell lines in this study,
and further verification of the efficacy of CAPE as a cancer
therapeutic for HSCC is required.
In conclusion, the present study provided initial
evidence that CAPE induces the mitochondria-mediated intrinsic
apoptosis pathway in HSCC by simultaneously regulating two major
members of the IAP protein family. These findings suggested that
CAPE offers the basis for a promising alternative strategy to cure
HSCC in clinical practice.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MSIT) (grant nos. 2019R1A2C1085896 and 2020R1C1C1005480).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding authors on reasonable
request.
Authors' contributions
SDC conceived and designed the study. HJK and MHA
performed all the experiments, designed the primers for RT-qPCR and
drafted the manuscript. JAS, SJC and HJY analyzed and interpreted
the data. HJK, MHA and SDC confirm the authenticity of all the raw
data. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Newman JR, Connolly TM, Illing EA, Kilgore
ML, Locher JL and Carroll WR: Survival trends in hypopharyngeal
cancer: A population-based review. Laryngoscope. 125:624–629. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eckel HE and Bradley PJ: Treatment options
for hypopharyngeal cancer. Adv Otorhinolaryngol. 83:47–53.
2019.PubMed/NCBI
|
|
3
|
Mura F, Bertino G, Occhini A and Benazzo
M: Surgical treatment of hypopharyngeal cancer: A review of the
literature and proposal for a decisional flow-chart. Acta
Otorhinolaryngol Ital. 33:299–306. 2013.PubMed/NCBI
|
|
4
|
Pandey P, Khan F, Upadhyay TK and Giri PP:
Therapeutic efficacy of caffeic acid phenethyl ester in cancer
therapy: An updated review. Chem Biol Drug Des. 102:201–216. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murtaza G, Karim S, Akram MR, Khan SA,
Azhar S, Mumtaz A and Bin Asad MH: Caffeic acid phenethyl ester and
therapeutic potentials. Biomed Res Int. 2014:1453422014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patel S: Emerging adjuvant therapy for
cancer: Propolis and its constituents. J Diet Suppl. 13:245–268.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kuo YY, Su LC, Chung CJ, Lin CY, Huo C,
Tseng JC, Huang SH, Lai CJ, Chen BC, Wang BJ, et al: Caffeic Acid
phenethyl ester is a potential therapeutic agent for oral cancer.
Int J Mol Sci. 16:10748–1066. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yu HJ, Shin JA and Cho SD: Inhibition of
focal adhesion kinase/paxillin axis by caffeic acid phenethyl ester
restrains aggressive behaviors of head and neck squamous cell
carcinoma in vitro. Arch Oral Biol. 146:1056112023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chung LC, Chiang KC, Feng TH, Chang KS,
Chuang ST, Chen YJ, Tsui KH, Lee JC and Juang HH: Caffeic acid
phenethyl ester upregulates N-myc downstream regulated gene 1 via
ERK pathway to inhibit human oral cancer cell growth in vitro and
in vivo. Mol Nutr Food Res. 612017.doi: 10.1002/mnfr.201600842.
|
|
10
|
Frassanito MA, Saltarella I, Vinella A,
Muzio LL, Pannone G, Fumarulo R, Vacca A and Mariggiò MA: Survivin
overexpression in head and neck squamous cell carcinomas as a new
therapeutic target (Review). Oncol Rep. 41:2615–2624.
2019.PubMed/NCBI
|
|
11
|
Tu H and Costa M: XIAP's profile in human
cancer. Biomolecules. 10:14932020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jaiswal PK, Goel A and Mittal RD:
Survivin: A molecular biomarker in cancer. Indian J Med Res.
141:389–397. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou LQ, Hu Y and Xiao HJ: The prognostic
significance of survivin expression in patients with HNSCC: A
systematic review and meta-analysis. BMC Cancer. 21:4242021.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Khan SA, Burke M, Zhu F, Yang DH, Dubyk C,
Mehra R, Lango MJ, Ridge JA, Sher DJ and Burtness B: Survivin
expression and impact on head and neck cancer outcomes. Oral Oncol.
112:1050492021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Obexer P and Ausserlechner MJ: X-linked
inhibitor of apoptosis protein-a critical death resistance
regulator and therapeutic target for personalized cancer therapy.
Front Oncol. 4:1972014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nagi C, Xiao GQ, Li G, Genden E and
Burstein DE: Immunohistochemical detection of X-linked inhibitor of
apoptosis in head and neck squamous cell carcinoma. Ann Diagn
Pathol. 11:402–406. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Ma X, Lu X, Cui L and Dong W:
Expression of inhibitor of apoptosis protein XIAP in laryngeal
carcinoma and its clinicopathological significance. Lin Chung Er Bi
Yan Hou Tou Jing Wai Ke Za Zhi. 21:973–975. 2007.(In Chinese).
PubMed/NCBI
|
|
18
|
Dohi T, Okada K, Xia F, Wilford CE, Samuel
T, Welsh K, Marusawa H, Zou H, Armstrong R, Matsuzawa S, et al: An
IAP-IAP complex inhibits apoptosis. J Biol Chem. 279:34087–34090.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Singh R, Letai A and Sarosiek K:
Regulation of apoptosis in health and disease: The balancing act of
BCL-2 family proteins. Nat Rev Mol Cell Biol. 20:175–193. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Park JS, Deng JH and Bai Y:
Cytochrome c oxidase subunit IV is essential for assembly and
respiratory function of the enzyme complex. J Bioenerg Biomembr.
38:283–291. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schwarzerová K, Bellinvia E, Martinek J,
Sikorová L, Dostál V, Libusová L, Bokvaj P, Fischer L, Schmit AC
and Nick P: Tubulin is actively exported from the nucleus through
the Exportin1/CRM1 pathway. Sci Rep. 9:57252019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang YL and Li XM: The IAP family:
Endogenous caspase inhibitors with multiple biological activities.
Cell Res. 10:169–177. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jo MJ, Jeong S, Yun HK, Kim DY, Kim BR,
Kim JL, Na YJ, Park SH, Jeong YA, Kim BG, et al: Genipin induces
mitochondrial dysfunction and apoptosis via downregulation of
Stat3/mcl-1 pathway in gastric cancer. BMC Cancer. 19:7392019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen M, Tong C, Wu Q, Zhong Z, He Q, Zeng
L and Xiao L: 6-Shogaol inhibits the cell migration of colon cancer
by suppressing the EMT process through the IKKβ/NF-κB/Snail
pathway. Integr Cancer Ther. 22:153473542311727322023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, Wei Y and Wei X: Napabucasin, a
novel inhibitor of STAT3, inhibits growth and synergises with
doxorubicin in diffuse large B-cell lymphoma. Cancer Lett.
491:146–161. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dong H, Hu L, Li W, Shi M, He L, Wang C,
Hu Y, Wang H, Wen C, Liu H and Yang X: Pyrimethamine inhibits cell
growth by inducing cell senescence and boosting CD8+
T-cell mediated cytotoxicity in colorectal cancer. Mol Biol Rep.
49:4281–4292. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Murugesan A, Lassalle-Claux G, Hogan L,
Vaillancourt E, Selka A, Luiker K, Kim MJ, Touaibia M and Reiman T:
Antimyeloma potential of caffeic acid phenethyl ester and its
analogues through Sp1 mediated downregulation of IKZF1-IRF4-MYC
axis. J Nat Prod. 83:3526–3535. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sari C, SÜmer C and Celep EyÜpoĞlu F:
Celep EyÜpoĞlu, Caffeic acid phenethyl ester induces apoptosis in
colorectal cancer cells via inhibition of survivin. Turk J Biol.
4:264–274. 2020. View Article : Google Scholar
|
|
30
|
Kleczka A, Kubina R, Dzik R, Jasik K,
Stojko J, Cholewa K and Kabała-Dzik A: Caffeic acid phenethyl ester
(CAPE) induced apoptosis in serous ovarian cancer OV7 cells by
deregulation of BCL2/BAX genes. Molecules. 25:35142020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Colombo D, Gatti L, Sjöstrand L, Carenini
N, Costantino M, Corna E, Arrighetti N, Zuccolo M, De Cesare M,
Linder S, et al: Caffeic acid phenethyl ester targets
ubiquitin-specific protease 8 and synergizes with cisplatin in
endometrioid ovarian carcinoma cells. Biochem Pharmacol.
197:1149002022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Marin EH, Paek H, Li M, Ban Y, Karaga MK,
Shashidharamurthy R and Wang X: Caffeic acid phenethyl ester exerts
apoptotic and oxidative stress on human multiple myeloma cells.
Invest New Drugs. 37:837–848. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pfeffer CM and Singh ATK: Apoptosis: A
target for anticancer therapy. Int J Mol Sci. 19:4482018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koff JL, Ramachandiran S and
Bernal-Mizrachi L: A time to kill: Targeting apoptosis in cancer.
Int J Mol Sci. 16:2942–2955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choi SJ, Ahn CH, Hong KO, Kim JH, Hong SD,
Shin JA and Cho SD: Molecular mechanism underlying the apoptotic
modulation by ethanol extract of Pseudolarix kaempferi in
mucoepidermoid carcinoma of the salivary glands. Cancer Cell Int.
21:4272021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chung TW, Choi H, Lee JM, Ha SH, Kwak CH,
Abekura F, Park JY, Chang YC, Ha KT, Cho SH, et al: Oldenlandia
diffusa suppresses metastatic potential through inhibiting matrix
metalloproteinase-9 and intercellular adhesion molecule-1
expression via p38 and ERK1/2 MAPK pathways and induces apoptosis
in human breast cancer MCF-7 cells. J Ethnopharmacol. 195:309–317.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheng AC, Jian CB, Huang YT, Lai CS, Hsu
PC and Pan MH: Induction of apoptosis by Uncaria tomentosa through
reactive oxygen species production, cytochrome c release, and
caspases activation in human leukemia cells. Food Chem Toxicol.
45:2206–2218. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Roy S and Nicholson DW: Cross-talk in cell
death signaling. J Exp Med. 192:F21–F25. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gavrilescu LC and Denkers EY: Apoptosis
and the balance of homeostatic and pathologic responses to
protozoan infection. Infect Immun. 71:6109–6115. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Suzuki S, Yamamoto M, Sanomachi T, Togashi
K, Sugai A, Seino S, Yoshioka T, Kitanaka C and Okada M:
Brexpiprazole, a serotonin-dopamine activity modulator, can
sensitize glioma stem cells to osimertinib, a third-generation
EGFR-TKI, via survivin reduction. Cancers. 11:9472019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sakoguchi-Okada N, Takahashi-Yanaga F,
Fukada K, Shiraishi F, Taba Y, Miwa Y, Morimoto S, Iida M and
Sasaguri T: Celecoxib inhibits the expression of survivin via the
suppression of promoter activity in human colon cancer cells.
Biochem Pharmacol. 73:1318–1329. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dean E, Jodrell D, Connolly K, Danson S,
Jolivet J, Durkin J, Morris S, Jowle D, Ward T, Cummings J, et al:
Phase I trial of AEG35156 administered as a 7-day and 3-day
continuous intravenous infusion in patients with advanced
refractory cancer. J Clin Oncol. 27:1660–1666. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yue C, Li RH, Chen C and Liu H: Study on
the relationship between XIAP gene and resistance of taxol in
ovarian cancer. Sichuan Da Xue Xue Bao Yi Xue Ban. 49:337–341.
2018.(In Chinese). PubMed/NCBI
|
|
45
|
Hehlgans S, Petraki C, Reichert S, Cordes
N, Rödel C and Rödel F: Double targeting of Survivin and XIAP
radiosensitizes 3D grown human colorectal tumor cells and decreases
migration. Radiother Oncol. 108:32–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Werner TA, Dizdar L, Nolten I, Riemer JC,
Mersch S, Schütte SC, Driemel C, Verde PE, Raba K, Topp SA, et al:
Survivin and XIAP-two potential biological targets in follicular
thyroid carcinoma. Sci Rep. 7:113832017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li Y, Gao W, Ma Y, Zhu G, Chen F and Qu H:
Dual targeting of survivin and X-linked inhibitor of apoptosis
protein suppresses the growth and promotes the apoptosis of gastric
cancer HGC-27 cells. Oncol Lett. 16:3489–3498. 2018.PubMed/NCBI
|
|
48
|
Fang W, Che X, Li G, Wang A, Wang Y, Shi
X, Hou K, Zhang X, Qu X and Liu Y: Sur-X, a novel peptide, kills
colorectal cancer cells by targeting survivin-XIAP complex. J Exp
Clin Cancer Res. 39:822020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liu TT, Yang H, Zhuo FF, Yang Z, Zhao MM,
Guo Q, Liu Y, Liu D, Zeng KW and Tu PF: Atypical E3 ligase ZFP91
promotes small-molecule-induced E2F2 transcription factor
degradation for cancer therapy. EBioMedicine. 86:1043532022.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dohi T, Xia F and Altieri DC: Altieri,
Compartmentalized phosphorylation of IAP by protein kinase A
regulates cytoprotection. Mol Cell. 27:17–28. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Arora V, Cheung HH, Plenchette S, Micali
OC, Liston P and Korneluk RG: Degradation of survivin by the
X-linked inhibitor of apoptosis (XIAP)-XAF1 complex. J Biol Chem.
282:26202–26209. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hoffman WH, Biade S, Zilfou JT, Chen J and
Murphy M: Transcriptional repression of the anti-apoptotic survivin
gene by wild type p53. J Biol Chem. 277:3247–3257. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mirza A, McGuirk M, Hockenberry TN, Wu Q,
Ashar H, Black S, Wen SF, Wang L, Kirschmeier P, Bishop WR, et al:
Human survivin is negatively regulated by wild-type p53 and
participates in p53-dependent apoptotic pathway. Oncogene.
21:2613–2622. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Baek HS, Kwon YJ, Ye DJ, Cho E, Kwon TU
and Chun YJ: CYP1B1 prevents proteasome-mediated XIAP degradation
by inducing PKCε activation and phosphorylation of XIAP. Biochim
Biophys Acta Mol Cell Res. 1866:1185532019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hong SW, Shin JS, Moon JH, Jung SA, Koh
DI, Ryu Y, Park YS, Kim DY, Park SS, Hong JK, et al:
Chemosensitivity to HM90822, a novel synthetic IAP antagonist, is
determined by p-AKT-inducible XIAP phosphorylation in human
pancreatic cancer cells. Invest New Drugs. 38:1696–1706. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chung TW, Moon SK, Chang YC, Ko JH, Lee
YC, Cho G, Kim SH, Kim JG and Kim CH: Novel and therapeutic effect
of caffeic acid and caffeic acid phenyl ester on hepatocarcinoma
cells: Complete regression of hepatoma growth and metastasis by
dual mechanism. FASEB J. 18:1670–1681. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Tang H, Yao X, Yao C, Zhao X, Zuo H and Li
Z: Anti-colon cancer effect of caffeic acid p-nitro-phenethyl ester
in vitro and in vivo and detection of its metabolites. Sci Rep.
7:75992017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kapałczyńska M, Kolenda T, Przybyła W,
Zajączkowska M, Teresiak A, Filas V, Ibbs M, Bliźniak R, Łuczewski
Ł and Lamperska K: 2D and 3D cell cultures-a comparison of
different types of cancer cell cultures. Arch Med Sci. 14:910–919.
2018.PubMed/NCBI
|
|
59
|
Mañós M, Giralt J, Rueda A, Cabrera J,
Martinez-Trufero J, Marruecos J, Lopez-Pousa A, Rodrigo JP, Castelo
B, Martínez-Galán J, et al: Multidisciplinary management of head
and neck cancer: First expert consensus using Delphi methodology
from the Spanish Society for Head and Neck Cancer (part 1). Oral
Oncol. 70:58–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kuo YY, Lin HP, Huo C, Su LC, Yang J,
Hsiao PH, Chiang HC, Chung CJ, Wang HD, Chang JY, et al: Caffeic
acid phenethyl ester suppresses proliferation and survival of TW2.6
human oral cancer cells via inhibition of Akt signaling. Int J Mol
Sci. 14:8801–8817. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Peng CY, Yang HW, Chu YH, Chang YC, Hsieh
MJ, Chou MY, Yeh KT, Lin YM, Yang SF and Lin CW: Caffeic Acid
phenethyl ester inhibits oral cancer cell metastasis by regulating
matrix metalloproteinase-2 and the mitogen-activated protein kinase
pathway. Evid Based Complement Alternat Med. 2012:7325782012.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Machiels JP, René Leemans C, Golusinski W,
Grau C, Licitra L and Gregoire V; EHNS Executive Board, ESMO
Guidelines Committee, : ESTRO Executive Board: Reprint of ‘Squamous
cell carcinoma of the oral cavity, larynx, oropharynx and
hypopharynx: EHNS-ESMO-ESTRO Clinical Practice Guidelines for
diagnosis, treatment and follow-up’. Ann Oncol. 31:1462–1475. 2020.
View Article : Google Scholar : PubMed/NCBI
|