Introduction
Lung cancer is responsible for the majority of
cancer-related deaths worldwide. The survival rate is related to
the stage of lung cancer, and there are regional differences
(1). Non-small cell lung cancer
(NSCLC) is the main type of lung cancer, which includes
adenocarcinoma, squamous cell carcinoma and large cell carcinoma,
accounting for 80–85% of the total number of lung cancer cases. Due
to the high malignancy of NSCLC, the irreversible distant
metastasis of tumor cells typically occurs prior to the initial
diagnosis in the majority of patients (2,3).
Therefore, novel therapeutic targets for the successful treatment
of lung cancer are urgently required.
Nuclear protein-1 (NUPR1), which is also known as
candidate of metastasis-1 (Com1), consists of 82 amino acids, and
its structure is highly unstable (4,5). NUPR1
is a transcriptional regulator that can be induced under several
cellular stresses, such as hypoxia and starvation (6–8),
thereby regulating autophagy through intracellular signaling to
counteract adverse situations (9).
First, during metastasis, NUPR1 facilitates the adaptation of tumor
cells to a new microenvironment and enables them to form new cancer
nests (10–12). Survival analysis of patients with
medullary thyroid tumors has indicated that NUPR1 is associated
with an unfavorable prognosis of patients with thyroid cancer. More
specifically, the expression level of NUPR1 is closely related to
lymph node metastasis, and 24.8% of patients with NUPR1 expression
have a higher recurrence rate (13,14).
Moreover, the downregulation of NUPR1 by siRNA has been
shown to significantly promote G1 phase cell cycle arrest (15,16)
and decrease the proliferation, in vitro soft agar colony
formation and tumorigenic ability of NSCLC cells in nude mice
(9). Similarly, a high expression
of NUPR1 also promotes the proliferation of pancreatic cancer cells
(17). Therefore, small compounds
or inhibitors that target NUPR1 for cancer therapy are
underdeveloped. For instance, chemicals that inhibit NUPR1 function
and mimic NUPR1 deficiency benefit the treatment of pancreatic
ductal adenocarcinoma (PDAC) (18).
Among these chemicals, trifluoperazine dihydrochloride (TFP) has
shown great promise to bind NUPR1 and induce cell growth arrest,
mimicking NUPR1 deficiency in pancreatic adenocarcinoma cells
(19); however, its primary
clinical use is as an antipsychotic drug for the treatment of
delusional schizophrenia, acute schizophrenic psychosis and chorea
(20). Its pharmacological
functions involve the blocking of dopamine receptors and
functioning as a calmodulin antagonist to bind to calmodulin and
interfere with Ca2+-calmodulin interactions (21). However, its efficacy in cancer
treatment has only been mentioned in a limited number of studies
reporting that TFP inhibits cell invasion and proliferation, and
induces cell death in several types of animal models and cancer
cell lines (22–25), without addressing the underlying
mechanisms or its potential targets.
The authors have previously demonstrated that the
downregulation of NUPR1 induces the formation of autophagic
vesicles and cell cycle arrest in NSCLC cells, inducing premature
senescence, thereby impairing the proliferation and colony
formation ability of NSCLC cells (9). This prompted the exploration of the
effects of manipulating NUPR1 by shRNA and small compounds
on lung cancer progression in vivo.
Adeno-associated virus (AAV)-based vectors lack
pathogenicity, but are efficient vehicles for gene delivery,
rendering them a potentially useful tool in gene therapy (26,27).
In this study, we used a recombinant AAV vector expressing
NUPR1 shRNA combined with TFP to examine the effect of a
NUPR1 ‘double kill’ approach on A549 cells in vitro and
in vivo. It was found that TFP combined with NUPR1
shRNA using AAV vector delivery enhanced the effects of TFP,
impairing autophagy, promoting premature senescence and inhibiting
tumor growth in vitro and in vivo. The findings of
this study may aid in the development of novel effective
therapeutic strategies for lung cancer.
Materials and methods
Cell lines
A549 and 293AD cells were obtained from the American
Type Culture Collection (CCL-185 and CRL-1573, respectively) and
were cultured according to the recommended protocols.
Viral particle production
Oligonucleotides encoding shRNA for NUPR1
(GGAGGACCCAGGACAGGATCC) or Firefly luciferase
(CGTACGCGGAATACTTCGATT) as a control were ligated into
pSUPER.retro.puro, and the fragment containing the H1 promoter and
hairpin sequences was subcloned into the pAAV vector (Cell Biolabs)
according to the manufacturer's protocol. The production and
purification of AAV were carried out as previously described
(28). The AAV titer was quantified
by qPCR analysis.
In vitro transgene expression
A549 cells were cultured in complete medium and
treated with AAV particles for 48 h. Fluorescence microscopy
(magnification, ×10; Olympus Corp.) was used to analyze the
infection efficiency of AAV particles. The infected cells were
collected for the analysis of the protein levels.
Western blot analysis
Whole cell protein extracts in 1X Laemmli buffer
(Bio-Rad, cat. no. 1610737) were resolved through 15% SDS-PAGE,
transferred to a nitrocellulose membrane, and probed with
antibodies against human NUPR1 (cat. no. 1:1,000; Abcam), SNAP25
(cat. no. ab109105; 1:1,000; Abcam), p62 (cat. no. 7695, 1:1,000;
Cell Signaling Technology), p16 (cat. no. ab108349, 1:2,000;
Abcam), p21 (cat. no. 9665; 1:2,000; Cell Signaling Technology),
p27 (cat. no. 9932; 1:2,000; Cell Signaling Technology) and
endogenous ACTB (cat. no. A-3853; 1:3,000; Sigma-Aldrich; Merck
KGaA) as a control for normalization. Peroxidase-conjugated
anti-mouse (cat. no. A-10654; 1:5,000; Thermo Fisher Scientific,
Inc.) and anti-rabbit IgG (cat. no. 31423; 1:5,000; Thermo Fisher
Scientific, Inc.) were used as secondary antibodies. The blots were
visualized on a Kodak X-ray film using an enhanced
chemiluminescence (ECL) detection substrate (cat. no. 32106, Thermo
Fisher Scientific, Inc.). ImageJ software (version 1.0; National
Institutes of Health) was used to quantify the intensities of band
signals, which were normalized to the ACTB internal controls.
Tumor xenografts
Animal care and surgical procedures were approved by
Tianjin Medical University and carried out in accordance with the
Institutional Animal Care and Use Committee guidelines (Tianjin
Medical University, permission no. SYXK: 2016-0012). A total of 16
male Fox Chase severe combined immunodeficient (SCID) mice were
housed at 18°C and supplied with water and laboratory chow. The
SCID mice were subjected to subcutaneous injections of A549 cells
(1.0×106 cells) in Matrigel (50:50) into the lower
flank. Ten days following tumor inoculation, the mice were randomly
divided into four groups and received tail vein injections of
AAV-iNUPR1, AAV-control (1×1011 vg/mouse), TFP, or
saline. To evaluate the effect of AAV and TFP treatment on tumor
growth, tumor volume was measured at the indicated time points
using a scaled ruler. After anesthetized by intraperitoneal
injection with 10% chloral hydrate (300 mg/kg body weight), the
mice were sacrificed by cervical dislocation. Death was confirmed
when the mice did not move, and then tumor tissues were collected
and weighed immediately, followed by fixation in buffered formalin
for histological and immunohistochemical analyses.
Transmission electron microscopy
Tumor tissue obtained from the animals was washed
with PBS and fixed with PBS containing 3% glutaraldehyde and 2%
paraformaldehyde (cat. nos. 340855 and P6148, respectively;
Sigma-Aldrich; Merck KGaA) (pH 7.3) for 2 h at room temperature.
Following fixation, the samples were washed with PBS and post-fixed
with 1% buffered osmium tetroxide (cat. no. 75632, Sigma-Aldrich;
Merck KGaA) for 30 min at room temperature and stained en bloc with
1% uranyl acetate (cat. no. 19481, Ted Pella, Inc.). The samples
were dehydrated in increasing concentrations of ethanol, embedded
in EMbed 812 medium (cat. no. 14120, Electron Microscopy Sciences)
and polymerized at 70°C for 2 days. Ultrathin sections were cut
using a Leica Ultracut microtome (Leica), stained with uranyl
acetate and lead citrate in a Leica EM Stainer and examined using a
JEM 1010 transmission electron microscope (JEOL) at an accelerating
voltage of 80 kV. Digital TEM images were acquired using an AMT
Imaging System (Advanced Microscopy Techniques).
Galactosidase β1 (GLB1) staining
Fresh tumor tissue was frozen in liquid nitrogen
following resection and processed for embedding in O.C.T. compound.
Tissue sections (5-µm-thick) were collected and processed for GLB1
staining. GLB1 staining was performed using a Senescence
β-Galactosidase Staining kit (cat. no. 9860, Cell Signaling
Technology) according to the manufacturer's protocol. Micrographs
were acquired under a light microscope (Nikon Eclipse Ti-U, Nikon
Instruments). Positive results for GLB1 staining were calculated
using Photoshop CS6 software (Adobe Systems).
Immunohistochemistry
Tumor tissue obtained from the animals was fixed in
formalin for 24 h at room temperature and incubated in 70% ethanol
for 48 h before being embedded in paraffin. The embedded tumors
were cut into 5-µm-thick sections, mounted on poly-L-lysine-coated
slides and then stained with hematoxylin and eosin (H&E) to
analyze the tumor histology. Immunohistochemical staining was
carried out as previously described (9). Cell proliferation was analyzed using
antibodies against Ki67 (Cell Signaling Technology, 1:400
dilution). Images were acquired with a CCD camera (Coolsnap ES,
Roper Scientific) equipped with Metamorph software (Molecular
Devices).
Statistical analysis
In this study, data were evaluated using SPSS
version 22.0 statistical software (IBM Corp.) and analyzed using an
unpaired two-tailed t-test or repeated measures analysis of
variance. Differences were considered statistically significant at
P<0.05.
Results
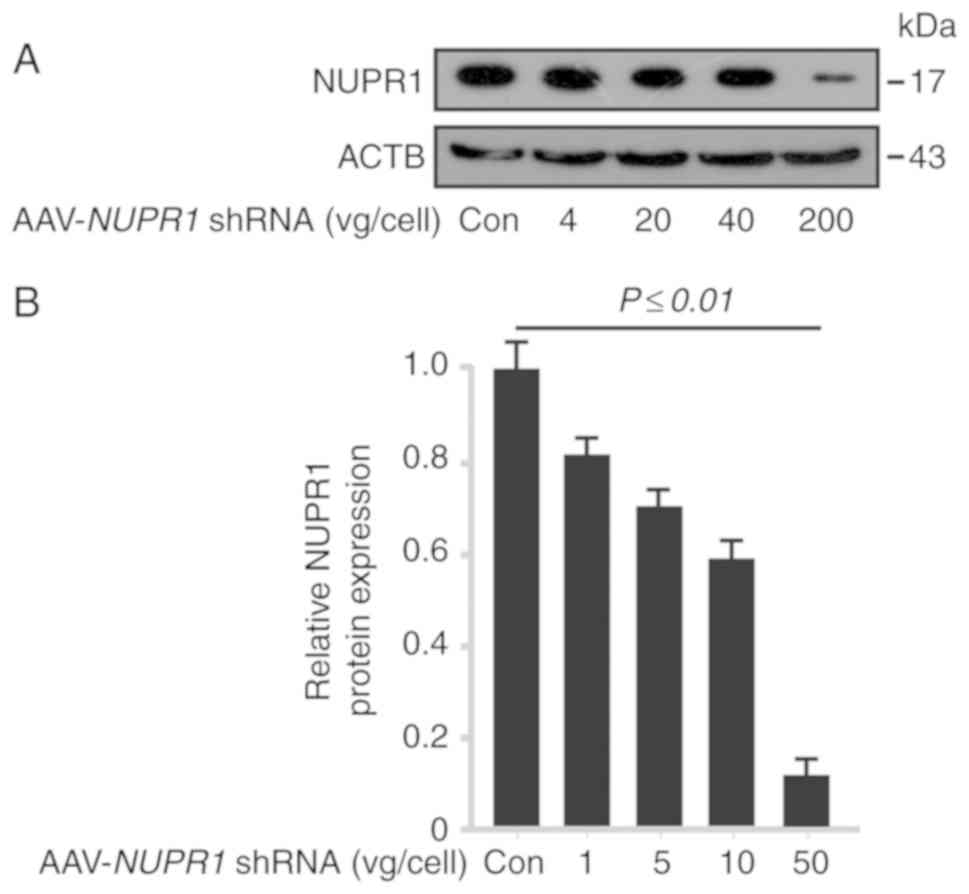
AAV-NUPR1 shRNA decreases NUPR1
expression levels in A549 cells in vitro
To verify the effects of AAV-NUPR1 shRNA on
the expression of NUPR1, AAV-control (cont), 4, 20, 40 and 200
vg/cell of the AAV-based vector containing NUPR1 shRNA were
used to infect A549 cells. Western blot analysis demonstrated that
AAV-NUPR1 shRNA decreased the NUPR1 protein expression
levels in A549 cells (Fig. 1A and
B) in a dose-dependent manner. These data indicated that
AAV-delivered shRNA significantly decreased NUPR1 protein
expression levels in vitro.
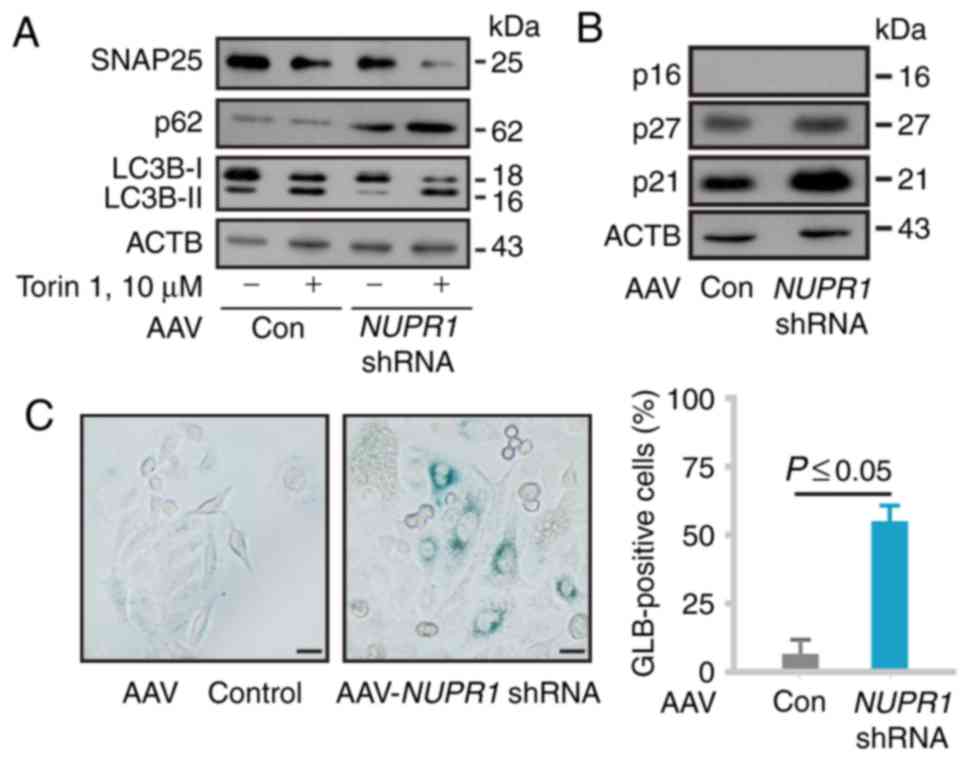
AAV-NUPR1 shRNA impairs the autophagic
process and causes premature senescence in A549 cells
Since it has already been verified that the
downregulation of NUPR1 impairs tumor cell autophagy (9), this study then examined the expression
of synaptosome associated protein 25 (SNAP25), sequestosome 1
(SQSTM1/p62) and microtubule-associated protein 1 light chain 3B
(MAP1LC3B/LC3B) in A549 cells by western blot analysis to confirm
its biological effects. As expected, NUPR1 depletion by the
AAV system decreased the SNAP25 protein expression level and
increased LC3B-I to LC3B-II conversion; all these effects were
enhanced by treatment with Torin 1 (an mTOR inhibitor and autophagy
inducer) (Fig. 2A), consistent with
previous findings. We then examined key cell cycle inhibitors (p16,
p21 and p27) by western blot analysis. AAV-mediated NUPR1
shRNA treatment resulted in the upregulation of p21 and p27
expression, but not p16 expression (Fig. 2B), which is a deleted mutation in
A549 cells. As expected, AAV-mediated NUPR1 depletion in
A549 cells led to a significant increase in the number of
GLB1-positive cells (Fig. 2C).
These data indicated that AAV-mediated NUPR1 shRNA impaired
tumor cell autophagy, leading to premature senescence in
vitro.
Combination of AAV-mediated NUPR1
knockdown with TFP decreases A549 tumorigenesis in vivo by a single
injection
TFP, an antipsychotic drug, has been shown to act as
a potent NUPR1 inhibitor, exerting antitumor effects (29). Subsequently, in this study, the
effect of AAV-mediated NUPR1 shRNA combined with TFP was
assessed on tumorigenesis in vivo, hypothesizing that TFP
enhances antitumor activity by NUPR1 depletion. A
subcutaneous model was employed to analyze the application
potential of TFP and AAV-NUPR1 shRNA. A549 cells were
subcutaneously injected into athymic nude mice. At 10 days
following tumor cell inoculation, all the cells formed visible
xenograft tumors. The mice were then randomly divided into four
groups and received tail vein injections of AAV-NUPR1 shRNA,
AAV-control (1×1011 vg/mouse), TFP, or saline as a
control. The tumor volumes in the AAV-NUPR1
shRNA/TFP-single-treated or double-treated groups were
significantly smaller than those in the control group, and the
tumor volumes of the double-treated group were the smallest among
all the groups (Fig. 3A).
Consistent with the tumor volume, the tumor weight and size in the
AAV-NUPR1 shRNA- and TFP-injected group were the most
reduced compared with the other groups (Fig. 3B and C). There was no significant
difference in tumor volume, weight and size between single therapy
and combination therapy; however, combination therapy still exerted
more beneficial tumor-suppressive effects, which was useful in
reducing the dose of the NUPR1 shRNA virus. Both the tumor
volume and weight were significantly decreased by combination
treatment (Fig. 3A-C).
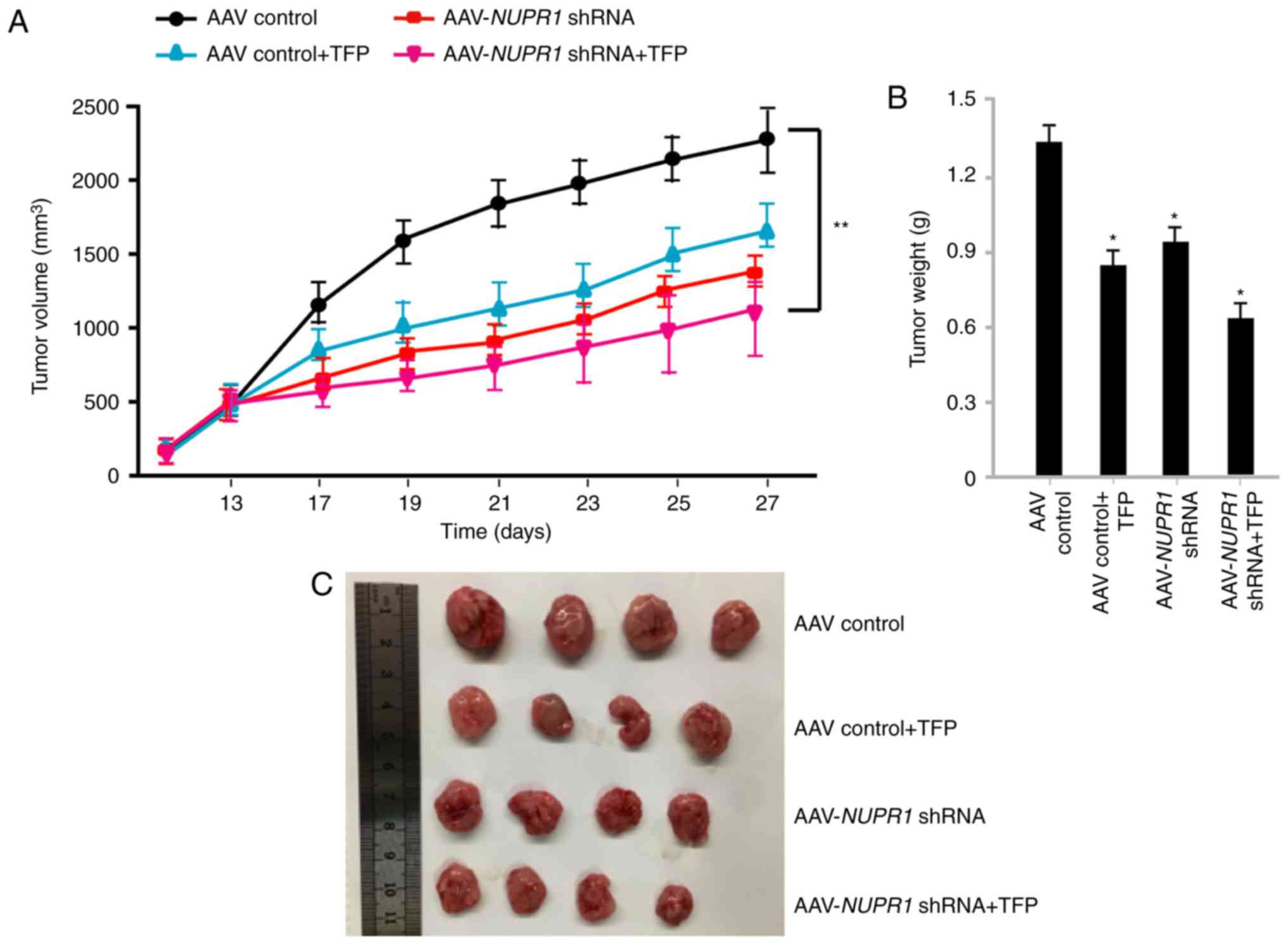 | Figure 3.AAV-NUPR1 shRNA and TFP
inhibits lung tumor growth. (A) Tumor growth in A549 tumor-bearing
mice that were injected with TFP, AAV-NUPR1 shRNA,
AAV-NUPR1 shRNA and TFP, or AAV-control particles (n=4). (B)
Tumors from mice treated with TFP, AAV-NUPR1 shRNA,
AAV-NUPR1 shRNA and TFP, or AAV-control particles were
weighed at the time of sacrifice (n=4). The data are presented as
the means ± SD of 4 mice. *P<0.05 and **P<0.01, compared to
the control group. (C) Images of tumors at the experimental
endpoint. NUPR1, nuclear protein-1; AAV, adeno-associated virus;
TFP, trifluoperazine dihydrochloride. |
AAV-mediated downregulation of NUPR1
and TFP treatment causes autophagy dysregulation and premature
senescence in vivo
To investigate whether the AAV-mediated
downregulation of NUPR1 has the same effect in vivo
as in vitro, histological examinations confirmed the
decrease in the tumor volume in the AAV-NUPR1 shRNA- and
TFP-injected groups, with a high cell death rate and substantially
decreased numbers of scattered tumor cells (Fig. 4A). Evidence to support this
phenomenon was also provided by transmission electron micrographs
exhibiting significantly increased autolysosomal vacuole formation
upon AAV-NUPR1 shRNA and TFP treatment on day 27 (Fig. 4B). This phenomenon that has been
previously described (9) is caused
by impaired autolysosomal processing, which is not favorable in
cancer cells. Immunohistochemical analyses confirmed that the NUPR1
expression level was decreased at 27 days following the injection
of AAV-NUPR1 shRNA or TFP, which was not observed in the
control tumors. Moreover, significant decreases in the NUPR1
expression level and in the number of cells positive for Ki67 were
observed, which is a marker of cell proliferation, in the
AAV-NUPR1 shRNA and TFP-treated groups compared with the
control group (Fig. 4C); no
significant difference was observed between single therapy and
combination therapy. Subsequently, whether AAV-NUPR1 shRNA
causes premature senescence in lung tumor xenografts was assessed.
As shown in the data summary (Fig.
4D), the AAV-NUPR1 shRNA-treated group and the combined
AAV-NUPR1 shRNA- and-TFP treated group exhibited
significantly more GLB1-stained cells than the other two groups,
suggesting that AAV-NUPR1 shRNA triggers premature
senescence in xenograft lung tumors. It is noteworthy that the
number of GLB-positive cells in the NUPR1 shRNA-treated
group was increased compared to that in the combination therapy
group. However, the tumor weight and size in the combination
therapy group were more greatly reduced compared with those in the
NUPR1 shRNA-treated group, suggesting that TFP may mediate
non-senescence cell death to impair lung tumor growth. The exact
function of TFP in tumors is not yet clear. The results of this
study indicated that it can interfere with the premature senescence
mediated by NUPR1 shRNA treatment; however, this warrants
further investigation. Overall, the results of this study indicated
that AAV-NUPR1 shRNA efficiently decreased A549 ×enograft
tumor cell proliferation, impaired autophagy, caused premature
senescence and further inhibited the growth of tumor xenografts in
nude mice; TFP enhanced all these effects, apart from premature
senescence. The combination of AAV-NUPR1 shRNA and TFP
treatment is effective in inhibiting the growth of xenograft
tumors.
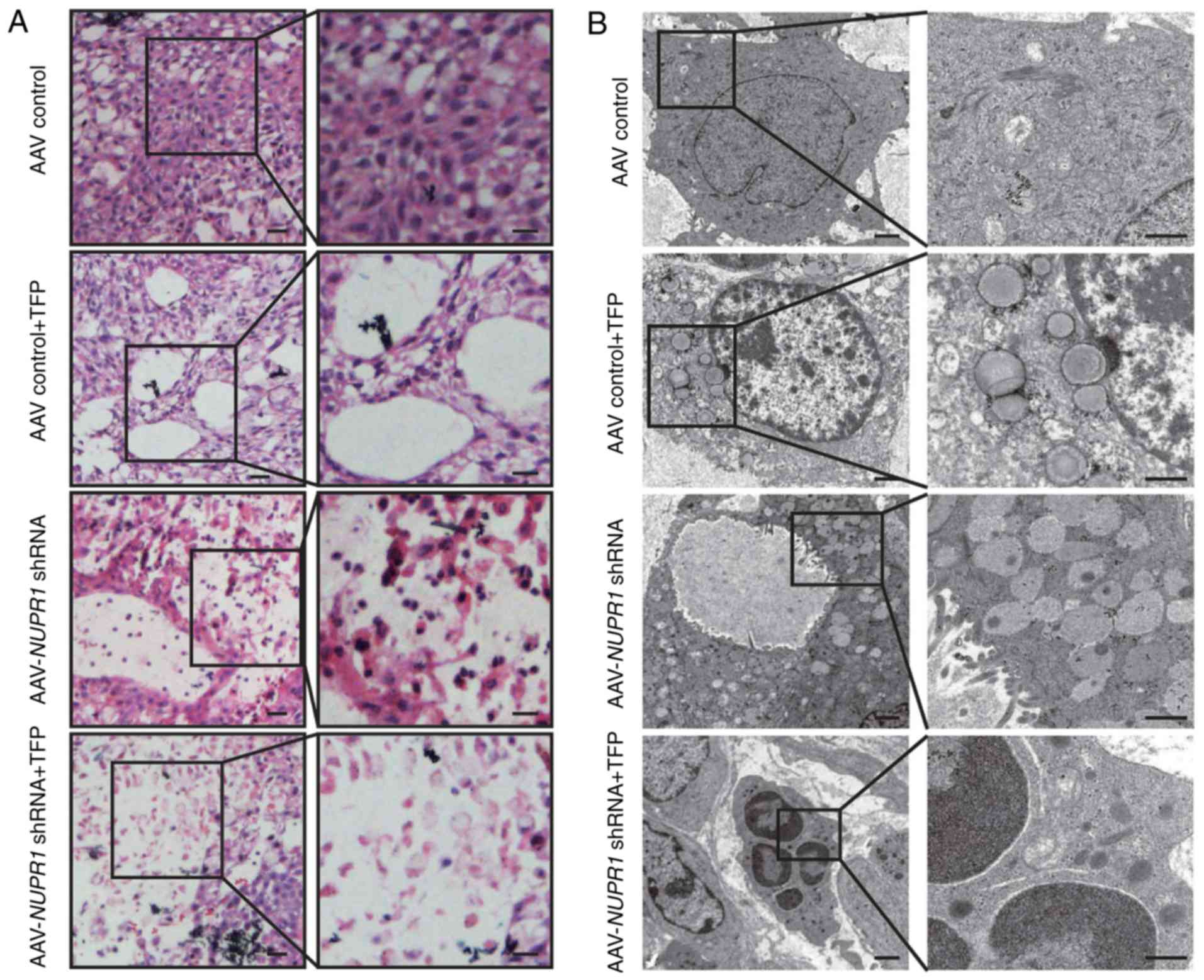 | Figure 4.Effects of AAV-NUPR1 shRNA and
TFP on A549 ×enograft tumor. (A) Representative transverse sections
of tumors from treated mice were stained with hematoxylin and eosin
(H&E). Scale bars, 100 µm. (B) A transmission electron
micrograph of tumor tissue excised at 27 days after administering
viral particles or a TFP injection. Scale bars, 200 nm. (C)
Sections of transplanted tumors infused with TFP, AAV-NUPR1
shRNA, AAV-NUPR1 shRNA and TFP, or AAV-control particles
were stained with NUPR1 and Ki67 antibodies (upper panels).
Intratumoral staining was quantified by the intensity multiplied by
the percentage of positive cells (mean ± SD) (lower panels).
*P<0.01, compared to the AAV-control. Scale bars, 10 µm. (D)
Representative images of GLB1 activity in frozen sections of
transplanted tumors infused with TFP, AAV-NUPR1 shRNA,
AAV-NUPR1 shRNA and TFP, or AAV-control particles (upper
panels). Quantification of GLB1-positive cells was conducted using
10 different fields of view from 3 independent experiments (mean ±
SEM) (lower panels). Scale bars, 50 µm. NUPR1, nuclear
protein-1; AAV, adeno-associated virus; TFP, trifluoperazine
dihydrochloride; GLB1, galactosidase β1. |
Discussion
As a transcriptional co-regulator, the expression
level of nuclear protein-1 (NUPR1) is relatively higher in the
majority of non-small cell lung cancer (NSCLC) cell lines examined
(9). In this study, it was found
that adeno-associated virus (AAV)-NUPR1 shRNA decreased
NUPR1 expression levels in A549 cells in a dose-dependent manner.
Moreover, NUPR1 knockdown also impaired the autophagic
process and caused premature senescence in A549 cells, consistent
with the results of previous studies by the authors (9). Both AAV-NUPR1 shRNA and
trifluoperazine dihydrochloride (TFP) treatments decreased A549
tumorigenesis in vivo, and combined treatment enhanced the
effects of single treatment using either AAV-NUPR1 shRNA or
TFP. The in vitro experimental data of the autophagic
process and premature senescence were reproduced in an in
vivo mouse model using A549 cell lines with NUPR1
knockdown by AAV-NUPR1 shRNA and/or treatment with TFP.
In a previous study by the authors, NUPR1
depletion impaired autophagic flux dynamics and induced
autolysosomal vacuolization and premature senescence via the
downregulation of SNAP25 expression (9). In addition, NUPR1 is a
stress-inducible transcriptional co-factor (30) that is specifically expressed in
several types of cancer (9,13,31)
and participates in a number of cancer-associated processes
(32–34). This suggests that NUPR1 plays an
important role in promoting cancer development and progression
without affecting, at least in part, adjacent normal tissues.
Consistent with these results, the data of this study suggest that
NUPR1 inhibition may be a novel therapeutic target for the
treatment of NSCLC.
The findings of this study demonstrated that
NUPR1 knockdown by the AAV system resulted in impaired tumor
cell autophagy, leading to premature senescence in vitro and
delayed xenograft tumor growth in vivo, consistent with the
results of a previous study by the authors using a lentiviral shRNA
delivery system (9). To the best of
our knowledge, this is the first report of the biological function
of NUPR1 knockdown by AAV-mediated shRNA delivery in NSCLC.
The data indicate that AAV is a very effective tool for shRNA
delivery and may be developed as a therapeutic strategy for
targeting NUPR1 in NSCLC. Although AAV is a potentially
useful tool in the field of gene therapy and is less pathogenic
than adenovirus (26), potential
risks caused by AAV integration remain (35). Another major concern associated with
the application of AAV-mediated NUPR1 shRNA delivery for the
treatment of tumors in vivo is its large-scale production,
since AAV production is time-consuming and costly. Therefore, it is
necessary to improve AAV-mediated NUPR1 shRNA therapy.
A number of clinical investigations have found that
antipsychotic drugs, such as penfluridol, chlorpromazine and
thioridazine may have potential anticancer potential effects for
clinical treatment (36). TFP,
which is a member of the phenothiazine class of antipsychotic
drugs, has been evaluated in phase I and phase II clinical trials
for the treatment of non-Hodgkin's lymphoma and glioblastoma
multiforme, respectively (37,38).
TFP activates the intrinsic apoptotic pathway in TNBC cells
(39) and A549 human lung cancer
cells (21). Recently, a
TFP-derived compound was produced and exhibited a dose-dependent
tumor regression with no neurological effects and an ability to
induce cell death mainly by necroptosis (29). Nevertheless, the present study
demonstrated that TFP binding indeed promotes changes in the
concentration and conformation of NUPR1, leading to beneficial
tumor-suppressive effects. Additionally, it was demonstrated that
the tumor growth inhibitory effect of AAV-mediated NUPR1
shRNA delivery combined with TFP treatment was more effective than
single treatment alone, although the difference was not highly
significant. Thus, TFP can reduce the therapeutic dose of the
NUPR1 shRNA virus. Reducing the dose of the NUPR1
shRNA virus may also aid in the prevention of adverse side-effects
such as transient increases in transaminases associated with
increased AAV capsid-specific T cells and decreased circulating
human factor IX levels. More importantly, the mice did not exhibit
significant weight loss or other signs of toxicity during
treatment, indicating the safety and efficiency of the combination
of AAV-mediated NUPR1 shRNA delivery with TFP as a strategy
for cancer therapy. It was also noted that the combination therapy
did not lead to the complete reduction of tumor growth and volume,
as this therapy targets NUPR1; as, it cannot be confirmed that all
lung cancer cells express and are dependent on NUPR1, the
effectiveness of the combination therapy is limited to
NUPR1-positive lung cancer cells. Thus, the combination of TFP with
AAV-mediated NUPR1 shRNA delivery may prolong
progression-free survival and achieve a better treatment response
in patients with NUPR1-positive tumors. Since TFP can bind NUPR1,
it may be predicted that radiolabeled TFP by fluorine-18 may be
used as a potential positron-emitting imaging agent, as well as an
antitumor drug for NUPR1-positive tumors.
In conclusion, to the best of our knowledge, this is
the first study on AAV-mediated NUPR1 shRNA delivery
combined with treatment with TFP for NUPR1-positive tumors. It was
demonstrated that the tail vein injection of AAV-mediated
NUPR1 shRNA with TFP significantly attenuated the growth of
lung cancer cell xenografts. These findings suggest that TFP
combined with AAV-mediated NUPR1 shRNA is a feasible and
potential antitumor approach for preclinical studies.
Acknowledgements
The authors would like to acknowledge the laboratory
equipment and platform support sponsored by the Tianjin Key
Laboratory of Medical Epigenetics, Key Laboratory of Immune
Microenvironment and Disease (Ministry of Education).
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 2018YFC1313002, 81825017,
81773034, 31600636, 81702245, 81874039, 81872350 and 81572271), the
Tianjin Municipal Science and Technology Commission (grant nos.
16JCYBJC24500, 18JCZDJC99100 and 19JCZDJC35600), and the open fund
of the State Key Laboratory of Medicinal Chemical Biology (Nankai
University) (grant no. 201501002).
Availability of data and materials
The datasets analyzed during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
ZL and ZM designed this study. YL, YY, JM, YS, RZ,
BC, YZ and XY performed the experiments. YL and JY analyzed the
data and prepared the figures. YL wrote the manuscript and XY
revised the manuscript. All the authors have read and approved the
final version of the manuscript for publication and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Animal care and surgical procedures were approved by
Tianjin Medical University and carried out in accordance with the
Institutional Animal Care and Use Committee guidelines (Tianjin
Medical University, permission no. SYXK: 2016-0012).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Herbst RS, Heymach JV and Lippman SM: Lung
cancer. N Engl J Med. 359:1367–1380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goruppi S, Patten RD, Force T and Kyriakis
JM: Helix-loop-helix protein p8, a transcriptional regulator
required for cardiomyocyte hypertrophy and cardiac fibroblast
matrix metalloprotease induction. Mol Cell Biol. 27:993–1006. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mallo GV, Fiedler F, Calvo EL, Ortiz EM,
Vasseur S, Keim V, Morisset J and Iovanna JL: Cloning and
expression of the rat p8 cDNA, a new gene activated in pancreas
during the acute phase of pancreatitis, pancreatic development, and
regeneration, and which promotes cellular growth. J Biol Chem.
272:32360–32369. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiang YF, Vaccaro MI, Fiedler F, Calvo EL
and Iovanna JL: Lipopolysaccharides induce p8 mRNA expression in
vivo and in vitro. Biochem Biophys Res Commun. 260:686–690. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taïeb D, Malicet C, Garcia S, Rocchi P,
Arnaud C, Dagorn JC, Iovanna JL and Vasseur S: Inactivation of
stress protein p8 increases murine carbon tetrachloride
hepatotoxicity via preserved CYP2E1 activity. Hepatology.
42:176–182. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zinke I, Schütz CS, Katzenberger JD, Bauer
M and Pankratz MJ: Nutrient control of gene expression in
Drosophila: Microarray analysis of starvation and sugar-dependent
response. EMBO J. 21:6162–6173. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mu Y, Yan X, Li D, Zhao D, Wang L, Wang X,
Gao D, Yang J, Zhang H, Li Y, et al: NUPR1 maintains autolysosomal
efflux by activating SNAP25 transcription in cancer cells.
Autophagy. 14:654–670. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jung SH, Lee A, Yim SH, Hu HJ, Choe C and
Chung YJ: Simultaneous copy number gains of NUPR1 and ERBB2
predicting poor prognosis in early-stage breast cancer. BMC Cancer.
12:3822012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iovanna JL: Expression of the
stress-associated protein p8 is a requisite for tumor development.
Int J Gastrointest Cancer. 31:89–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vasseur S, Hoffmeister A, Garcia S, Bagnis
C, Dagorn JC and Iovanna JL: p8 is critical for tumour development
induced by rasV12 mutated protein and E1A oncogene. EMBO Rep.
3:165–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ito Y, Yoshida H, Motoo Y, Iovanna JL,
Tomoda C, Uruno T, Takamura Y, Miya A, Kobayashi K, Matsuzuka F, et
al: Expression of p8 protein in medullary thyroid carcinoma.
Anticancer Res. 25:3419–3423. 2005.PubMed/NCBI
|
|
14
|
Su SB, Motoo Y, Iovanna JL, Xie MJ and
Sawabu N: Effect of camostat mesilate on the expression of
pancreatitis-associated protein (PAP), p8, and cytokines in rat
spontaneous chronic pancreatitis. Pancreas. 23:134–140. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo X, Wang W, Hu J, Feng K, Pan Y, Zhang
L and Feng Y: Lentivirus-mediated RNAi knockdown of NUPR1 inhibits
human nonsmall cell lung cancer growth in vitro and in vivo. Anat
Rec (Hoboken). 295:2114–2121. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vasseur S, Vidal Mallo G, Fiedler F,
Bödeker H, Cánepa E, Moreno S and Iovanna JL: Cloning and
expression of the human p8, a nuclear protein with mitogenic
activity. Eur J Biochem. 259:670–675. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hamidi T, Algül H, Cano CE, Sandi MJ,
Molejon MI, Riemann M, Calvo EL, Lomberk G, Dagorn JC, Weih F, et
al: Nuclear protein 1 promotes pancreatic cancer development and
protects cells from stress by inhibiting apoptosis. J Clin Invest.
122:2092–2103. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Santofimia-Castaño P, Rizzuti B, Abián O,
Velázquez-Campoy A, Iovanna JL and Neira JL: Amphipathic helical
peptides hamper protein-protein interactions of the intrinsically
disordered chromatin nuclear protein 1 (NUPR1). Biochim Biophys
Acta Gen Subj. 1862:1283–1295. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Neira JL, Bintz J, Arruebo M, Rizzuti B,
Bonacci T, Vega S, Lanas A, Velázquez-Campoy A, Iovanna JL and
Abián O: Identification of a drug targeting an intrinsically
disordered protein involved in pancreatic adenocarcinoma. Sci Rep.
7:397322017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang L, Yu J, Pan H, Hu P, Hao Y, Cai W,
Zhu H, Yu AD, Xie X, Ma D and Yuan J: Small molecule regulators of
autophagy identified by an image-based high-throughput screen. Proc
Natl Acad Sci USA. 104:19023–19028. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen QY, Wu LJ, Wu YQ, Lu GH, Jiang ZY,
Zhan JW, Jie Y and Zhou JY: Molecular mechanism of trifluoperazine
induces apoptosis in human A549 lung adenocarcinoma cell lines. Mol
Med Rep. 2:811–817. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Annabi B, Pilorget A, Bousquet-Gagnon N,
Gingras D and Béliveau R: Calmodulin inhibitors trigger the
proteolytic processing of membrane type-1 matrix metalloproteinase,
but not its shedding in glioblastoma cells. Biochem J. 359:325–333.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen MH, Lin KJ, Yang WLR, Kao YW, Chen
TW, Chao SC, Chang PM, Liu CY, Tzeng CH, Chao Y, et al: Gene
expression-based chemical genomics identifies heat-shock protein 90
inhibitors as potential therapeutic drugs in cholangiocarcinoma.
Cancer. 119:293–303. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shin SY, Kim CG, Hong DD, Kim JH and Lee
YH: Implication of Egr-1 in trifluoperazine-induced growth
inhibition in human U87MG glioma cells. Exp Mol Med. 36:380–386.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yeh CT, Wu ATH, Chang PMH, Chen KY, Yang
CN, Yang SC, Ho CC, Chen CC, Kuo YL, Lee PY, et al:
Trifluoperazine, an antipsychotic agent, inhibits cancer stem cell
growth and overcomes drug resistance of lung cancer. Am J Respir
Crit Care Med. 186:1180–1188. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miller N: Glybera and the future of gene
therapy in the European Union. Nat Rev Drug Discov. 11:4192012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi J, Zheng D, Liu Y, Sham MH, Tam P,
Farzaneh F and Xu R: Overexpression of soluble TRAIL induces
apoptosis in human lung adenocarcinoma and inhibits growth of tumor
xenografts in nude mice. Cancer Res. 65:1687–1692. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiao W, Chirmule N, Schnell MA, Tazelaar
J, Hughes JV and Wilson JM: Route of administration determines
induction of T-cell-independent humoral responses to
adeno-associated virus vectors. Mol Ther. 1:323–329. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Santofimia-Castaño P, Xia Y, Lan W, Zhou
Z, Huang C, Peng L, Soubeyran P, Velázquez-Campoy A, Abián O,
Rizzuti B, et al: Ligand-based design identifies a potent NUPR1
inhibitor exerting anticancer activity via necroptosis. J Clin
Invest. 129:2500–2513. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goruppi S and Iovanna JL: Stress-inducible
protein p8 is involved in several physiological and pathological
processes. J Biol Chem. 285:1577–1581. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li J, Ren S, Liu Y, Lian Z, Dong B, Yao Y
and Xu Y: Knockdown of NUPR1 inhibits the proliferation of
glioblastoma cells via ERK1/2, p38 MAPK and caspase-3. J
Neurooncol. 132:15–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gironella M, Malicet C, Cano C, Sandi MJ,
Hamidi T, Tauil RMN, Baston M, Valaco P, Moreno S, Lopez F, et al:
p8/nupr1 regulates DNA-repair activity after double-strand gamma
irradiation-induced DNA damage. J Cell Physiol. 221:594–602. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Malicet C, Dagorn JC, Neira JL and Iovanna
JL: p8 and prothymosin alpha: Unity is strength. Cell Cycle.
5:829–830. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sandi MJ, Hamidi T, Malicet C, Cano C,
Loncle C, Pierres A, Dagorn JC and Iovanna JL: p8 expression
controls pancreatic cancer cell migration, invasion, adhesion, and
tumorigenesis. J Cell Physiol. 226:3442–3451. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chandler RJ, Sands MS and Venditti CP:
Recombinant adeno- associated viral integration and genotoxicity:
Insights from animal models. Hum Gene Ther. 28:314–322. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu L, Liu YY, Li ZX, Zhao Q, Wang X, Yu Y,
Wang YY, Wang YQ and Luo F: Anti-tumor effects of penfluridol
through dysregulation of cholesterol homeostasis. Asian Pac J
Cancer Prev. 15:489–494. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hait WN, Byrne TN, Piepmeier J, Durivage
HJ, Choudhury S, Davis CA and Gates JA: The effect of calmodulin
inhibitors with bleomycin on the treatment of patients with high
grade gliomas. Cancer Res. 50:6636–6640. 1990.PubMed/NCBI
|
|
38
|
Hait WN, Morris S, Lazo JS, Figlin RJ,
Durivage HJ, White K and Schwartz PE: Phase I trial of combined
therapy with bleomycin and the calmodulin antagonist,
trifluoperazine. Cancer Chemother Pharmacol. 23:358–362. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Feng Z, Xia Y, Gao T, Xu F, Lei Q, Peng C,
Yang Y, Xue Q, Hu X, Wang Q, et al: The antipsychotic agent
trifluoperazine hydrochloride suppresses triple-negative breast
cancer tumor growth and brain metastasis by inducing G0/G1 arrest
and apoptosis. Cell Death Dis. 9:10062018. View Article : Google Scholar : PubMed/NCBI
|