Introduction
Cardiovascular disease has a high incidence in
numerous countries and is one of the most common threats to human
health, accounting for 30% of all deaths (1). Atherosclerosis (AS) is the basis of
various cardiovascular diseases. Numerous studies have demonstrated
that vascular endothelial injury is the initial step of AS
development (2,3). Oxidative stress can lead to the
imbalance in intracellular antioxidant capacity, thus producing a
large quantity of reactive oxygen species (ROS), inducing lipid
peroxidation and biomacromolecular degeneration, which is
considered to be the main factor leading to vascular endothelial
cell injury (4). Therefore, the
study of endothelial anti-oxidative stress injury and the
inhibition of endothelial apoptosis is of great importance for the
treatment of cardiovascular and cerebrovascular diseases.
The dried root-tuber of Ophiopogon japonicus
has been historically used as a common agent in the clinical
treatment of cardiovascular and cerebrovascular diseases (5). Radix Ophiopogonis is often used
together with ginseng and Schisandra chinensis, and is an
important raw material in traditional Chinese medicine in the form
of shengmai powder, and application of shengmai injection and
shenmai injection (6,7). The results of previous experiments
and clinical studies indicate that Radix Ophiopogonis and its
preparations have significant effects on the cardiovascular system,
and can improve myocardial contractility, enhance cardiac blood
output, and reduce cardiac load and myocardial oxygen consumption
(8–10). In clinic, Radix Ophiopogonis has a
notable effect on chronic cardiac insufficiency and coronary heart
disease (11). Pharmacological
experiments have demonstrated that Maidong injection protects
against myocardial ischemia, and methylophiopogonanone B (MO-B) is
one of the effective substances of Maidong injection (6,12).
Using a male rabbit model of anterior descending coronary artery
ligation to observe the influence of Maidong injection on
microstructures in experimental myocardial infarction and
myocardial ischemia, it was observed that the Maidong injection
group exhibited an increased negative rate of myocardial damage
than the control group; the incidence of myocardial infarction was
lower compared with the control group (13). At present, the majority of studies
on the main therapeutic substances of Radix Ophiopogonis are
focused on saponins, while few are focused on
homoisoflavonoids.
The MO-B of Radix Ophiopogonis is a major
homoisoflavonoid monomer isolated from Ophiopogon japonicus.
A recent study reported multiple activities of MO-B in various
systems, with the highest antioxidant activity being exhibited
in vitro (14). MO-B
promotes Rho activation and tubulin depolymerisation (15), and inhibits hypoxia-inducible
factor (HIF)-1 activity (16).
Furthermore, Ito et al (17) reported that MO-B can inhibit
melanosome transfer in normal human epidermal melanocytes. It has
also been reported that MO-B exerts significant anti-tumor
activities against HeLa cells (18); however, despite the various
biological activities of MO-B, the cellular function of MO-B in the
prevention of cardiovascular diseases in human umbilical vein
endothelial cells (HUVECs), and its underlying molecular mechanism
remain unknown. Thus, the present study investigated the effects of
MO-B against injury on H2O2-exposed HUVECs in
order to provide experimental evidence for its potential clinical
use in the treatment of cardiovascular diseases.
Our study demonstrated that MO-B prevents HUVECs
from H2O2-induced apoptosis by modulating
nicotinamide adeninde dinucleotide phosphate (NADPH) signaling,
caspase-3 and Bcl-2/(Bcl-2-associated X protein (Bax), indicating
that MO-B could be a potential agent in promoting the viability of
endothelial cells.
Materials and methods
Materials, reagents and
antibodies
Ophiopogon japonicus was obtained from farms
in Cixi (Zhejiang, China). MO-B was extracted from Ophiopogon
japonicus using high-speed counter-current chromatography
(19) and the yield was ~0.2–0.4
mg/g in tuber roots of Ophiopogon japonicus.
High-performance liquid chromatography (HPLC) was conducted to
measure the purity of MO-B, which was >97% (Fig. S1).HPLC was performed using a
Shimadzu C18 column (5 µm 250×4.6 mm). The volume ratio of mobile
solvents A (water) and B (acetonitrile) was maintained at 35:65,
and the temperature was set at 30°C. The flow rate of the mobile
phase was 1 ml/min. The detection wavelength was 285 nm. MO-B was
then dissolved to 10, 20, 40 and 50 µM in dimethyl sulfoxide for
cell treatment.
Antibodies targeting Bax (ab182733), Bcl-2
(ab182858), cleaved caspase-3 (ab32042), neutrophil cytochrome
b light chain (p22phox; ab80896) and GAPDH (ab9482), goat
anti-mouse horseradish peroxidase IgG (ab6789) and goat anti-rabbit
IgG horseradish peroxidase (ab6721) secondary antibodies, were
purchased from Abcam. Cell Counting Kit-8 (CCK-8), ROS and
malondialdehyde (MDA) detection kits, radioimmunoprecipitation
assay (RIPA) lysis buffer, a BCA Protein Assay kit and superoxide
dismutase (SOD) assay kit with WST-8 were purchased from Beyotime
Institute of Biotechnology.
Cell culture
HUVECs were obtained from Procell Life Science &
Technology Co., Ltd. and the STR validation of the cell line was
performed by the company, which revealed no cross contamination of
human cells. HUVECs were grown in Ham's F-12 K medium with 0.1
mg/ml Heparin, 0.03–0.05 mg/ml Endothelial Cell Growth Supplement,
10% fetal bovine serum and 1% penicillin/streptomycin (Procell Life
Science & Technology Co., Ltd.), and maintained at 37°C in a
humidified incubator with 5% CO2. The cells were
sub-cultured every 2–3 days with 0.25% trypsin digestion. Cells
between passages 5–12 were used for the subsequent the
experiments.
Cell viability assay
To evaluate cell viability, cells were enzymatically
harvested as aforementioned, counted in a hemocytometer and
sub-cultured in 96-well plates at a density of 5×103
cell/well. HUVECs cultivated for 24 h in medium at 37°C without
(control) or with MO-B (10, 20, 40 and 50 µM) were incubated with
H2O2 (1,000 µM) for 60 min. Finally, the
medium was discarded and 100 µl fresh medium containing 10% CCK-8
agent was added to each well for incubation for 1 h at 37°C. The
absorbance at 450 nm was measured using a Varioskan Flash reader
(Thermo Fisher Scientific, Inc.).
MDA and SOD assays
HUVECs were cultured at a density of
2×105 cells/well in 6-well plates and cultured overnight
at 37°C before being treated for 24 h without (control) or with
MO-B (10, 20, 40 and 50 µM), and then stimulated with
H2O2 (1,000 µM) for 6 h. The aforementioned
assay kits (Beyotime Institute of Biotechnology) were then used to
measure the MDA levels and SOD activity, respectively, according to
the manufacturer's protocols.
Intracellular ROS quantification
The level of intracellular ROS was determined by the
change in fluorescence emission of the fluorescent probe
2′,7′-dichlorofluorescein diacetate (DCFH-DA). Briefly,
2×105 HUVECs were cultured into 6-well plates at 37°C
and treated with the indicated concentrations of MO-B (10, 40 and
50 µM) for 24 h, followed by treatment with
H2O2 (1,000 µM) for 1 h. Cells were
trypsinized and washed with PBS, and then incubated with 10 mmol/l
DCFH-DA for 30 min at 37°C. Subsequently, cells were washed with
PBS twice and analyzed using a BD ACCURIC6 PLUS flow cytometer and
BD Accuri C6 software (BD Biosciences).
Analysis of apoptosis
After treatment with or without MO-B (10, 20 and 40
µM) for 24 h, cells were exposed to 1,000 µM
H2O2 for 6 h and then washed with ice-cold
PBS. Apoptosis was analyzed using an Annexin V-fluorescein
isothiocyanate/propidium iodide Apoptosis Detection Kit (Beyotime
Institute of Biotechnology). Cells were stained according to the
manufacturer's instructions. The proportion of apoptotic cells in
1×105 labeled cells was quantified using a BD Accuri C6
Plus flow cytometry and BD Accuri C6 software (BD Biosciences).
Reverse transcription-quantitative PCR
(RT-qPCR)
Cells were pretreated at 37°C without (control) or
with MO-B (10, 20 and 50 µM) for 24 h and then exposed to 1,000 µM
H2O2 for 6 h. The cells were then used for
RNA extraction.
Total RNA extraction was performed using the RNAiso
Plus reagent (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocols. The concentration of RNA was determined
by measuring the absorbance at 260 and 320 nm using a Nanodrop 2000
(Thermo Fisher Scientific, Inc.). Complementary DNA was generated
from 500 ng total RNA using Super Script II Reverse Transcriptase
(Takara Biotechnology Co., Ltd.), according to the manufacturer's
protocol. The reverse transcription reaction was as follows: 37°C
for 30 min and 85°C for 5 min. qPCR analysis was performed using
SYBR® GREEN PCR master mix in a reaction volume of 20 µl
using a 7500 Fast Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The annealing temperature used was 60°C
for 30 sec. Relative gene expression was calculated by the
2−ΔΔCq method (20),
and the values were normalized to the endogenous reference β-actin.
Primer sequences were as follows (forward, 5′-3′ and reverse,
5′-3′): p22phox, CAGTGGTACTTTGGTGCCTACTCCandGGTGGAGCCCTTCTTCCTCT;
Bcl-2, CGACGACTTCTCCCGCCGCTACCGC and CCGCATGCTGGGGCCGTACAGTTCC;
Bax: TCCACCAAGAAGCTGAGCGAG and GTCCAGCCCATGATGGTTCT; caspase-3,
AATTGTGGAATTGATGCGTGATGT and ATAATAACCAGGTGCTGTGGAGTA; β-actin,
GTGGGGCGCCCCAGGCACC and CTCCTTAATGTCACGCACGATTTC.
Western blot analysis
Cells were pretreated with MO-B (10, 20, 40 and 50
µM) for 24 h and then exposed to 1,000 µM
H2O2 for 6 h. Subsequently, the cells were
collected and lysed in RIPA lysis buffer containing 2 mM PMSF, and
the protein concentration in cell lysates was determined by BCA
assay. Cell lysates containing 60 µg total protein were subjected
to 12% SDS-PAGE (Bis-Tris Midi-Gels; Thermo Fisher Scientific,
Inc.) and then transferred to PVDF membranes. The membranes were
blocked in TBS with 0.1% Tween-20 (TBST) containing 1% BSA at 4°C
overnight, and then incubated with primary antibodies (1:5,000) in
TBST at room temperature for 2 h. The membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies (1:2,000) in
TBST at room temperature for 2 h. Proteins were visualized using an
ECL kit (Beyotime Institute of Biotechnology) and detected using a
Chemi Doc XRS imaging system (Bio-Rad Laboratories). GAPDH was used
as a loading control.
Statistical analysis
Data are expressed as the mean ± standard error from
three independent experiments. Statistical analysis was performed
with statistical software SPSS 18.0 (SPSS, Inc.). Data were
analyzed by one-way analysis of variance followed by the Least
Significant Difference test. P<0.05 was considered to indicate a
statistically significant difference.
Results
MO-B protects HUVECs against
H2O2-induced cell death
To determine the protective effects of MO-B on
HUVECs under H2O2 stress, a CCK-8 assay was
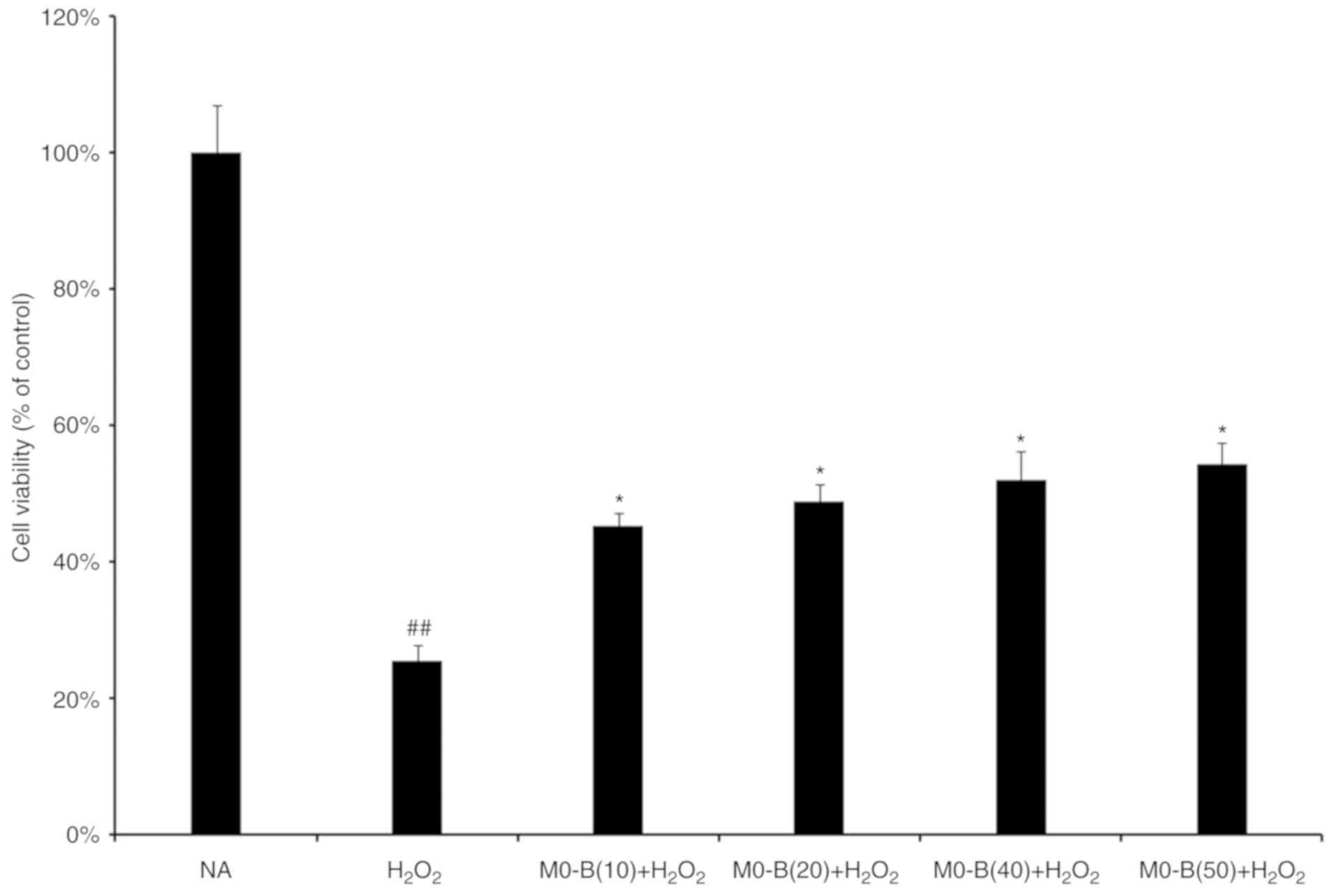
performed. As presented in Fig. 1,
H2O2 treatment for 24 h significantly
inhibited HUVEC viability compared with in untreated cells.
However, pretreatment of cells with MO-B for 24 h at concentrations
of 10, 20, 40 and 50 µm significantly ameliorated the effects of
H2O2 on cytotoxicity compared with
H2O2 treatment alone; cell activity increased
by 30% under 50 µM MO-B treatment (Fig. 1). This suggested that MO-B could
protect endothelial cells against
H2O2-induced cell death in a
concentration-dependent manner.
MO-B attenuates
H2O2-induced oxidative stress in HUVECs
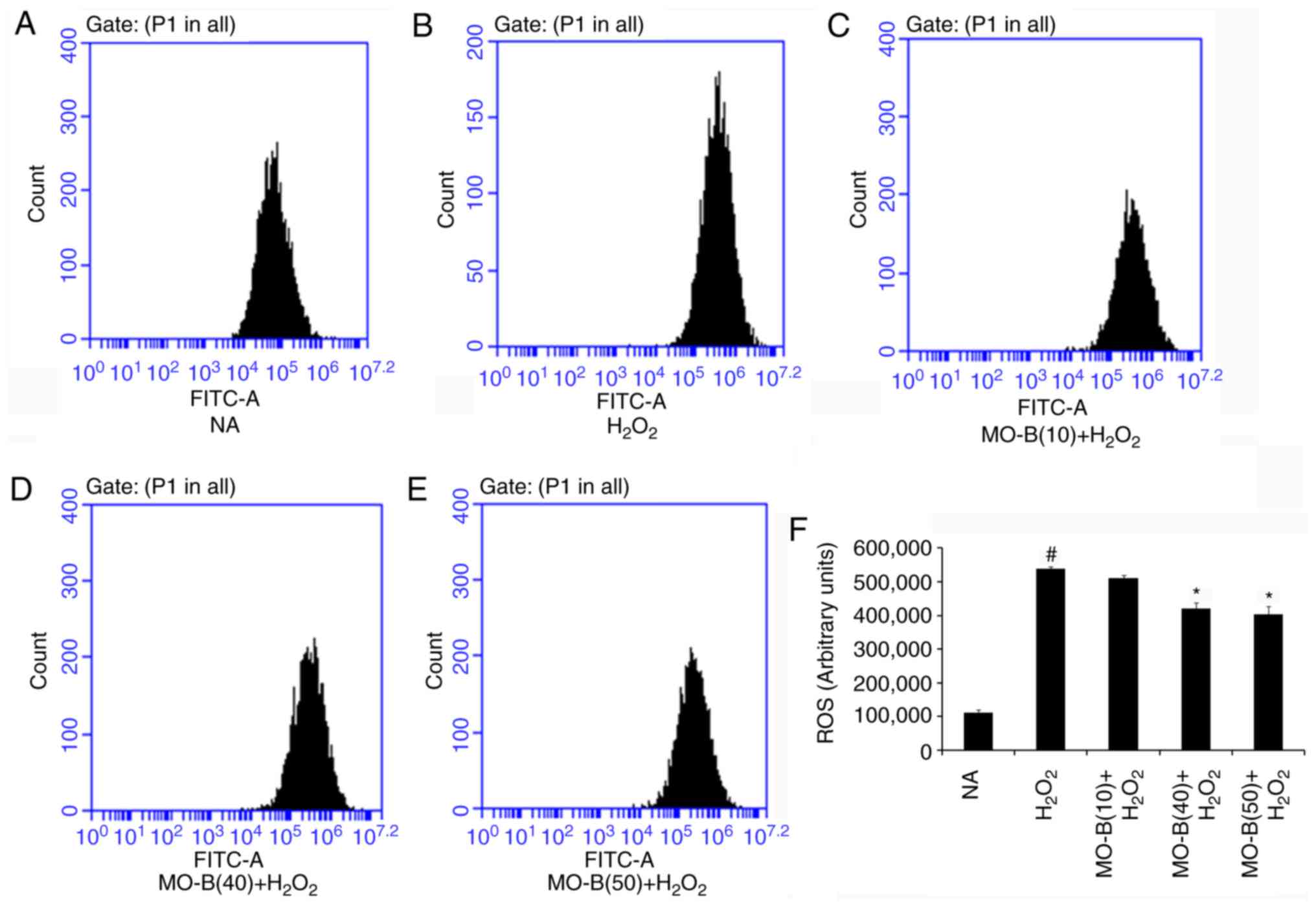
To further examine whether MO-B could protect HUVECs
via an antioxidant mechanism, the intracellular production of ROS
and MDA, and SOD activity were investigated. As presented in
Fig. 2, intracellular ROS levels
were significantly increased in the
H2O2-treated group compared with the control
group, while 40 and 50 µM MO-B significantly reduced ROS levels
compared with H2O2 treatment alone. This
indicated that H2O2 exerts its cytotoxicity
via oxidative injury and MO-B can mitigate such injury. Similarly,
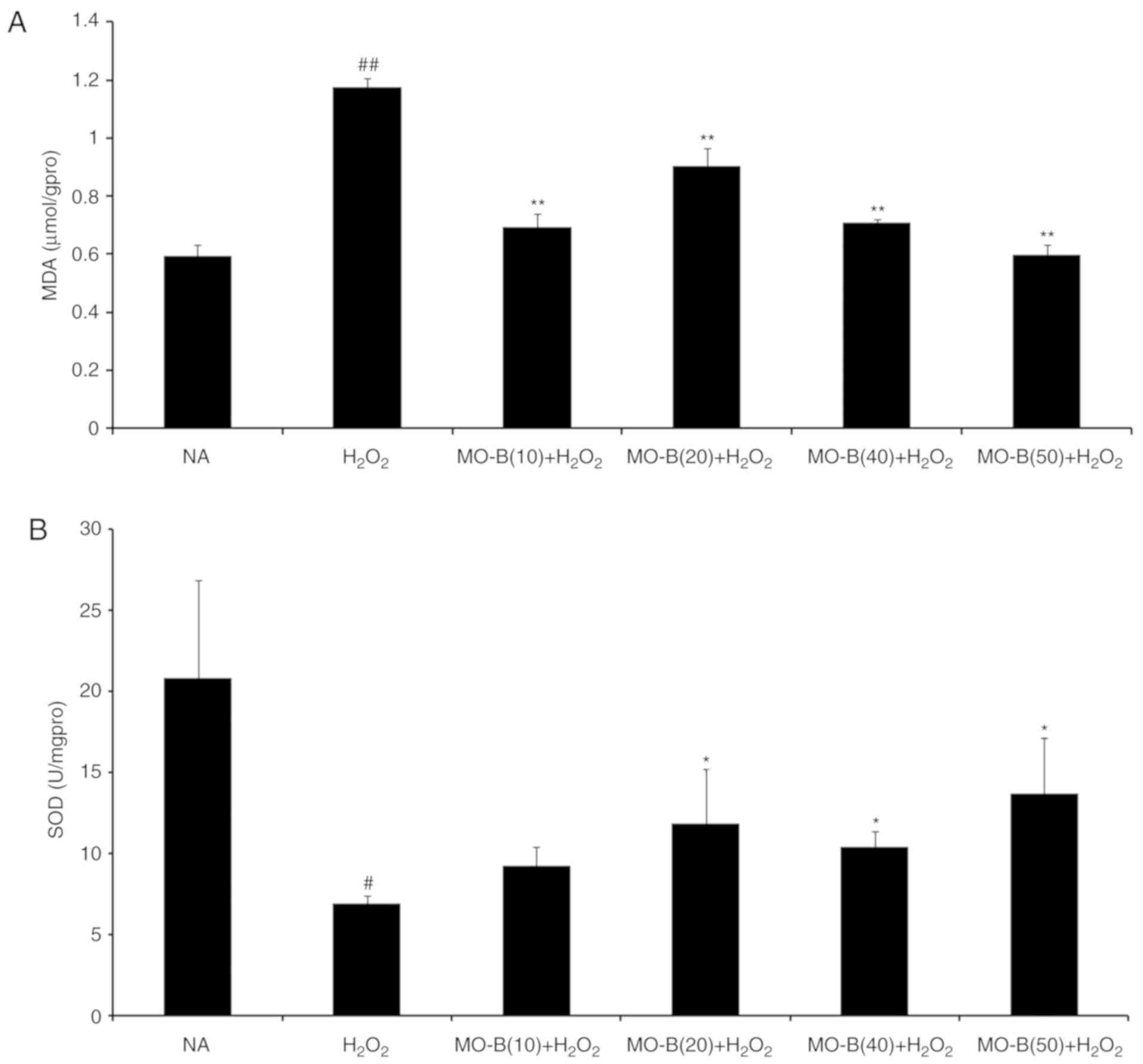
the levels of MDA, which is an indicator of lipid peroxidation
(21), were significantly
increased under H2O2 treatment compared with
the control, but decreased with MO-B pretreatment compared with
H2O2 treatment alone (Fig. 3A). This suggested that the
increased ROS and MDA produced are scavenged by MO-B, indicating
its antioxidative properties. Subsequently, the antioxidant enzyme
SOD was examined. SOD is a type of superoxide free radical
scavenger that naturally occurs in living organisms, and is an
active substance capable of eliminating harmful substances produced
during metabolic processes (22).
In our study, SOD activity was significantly decreased in HUVECs
treated with H2O2 for 6 h compared with
untreated cells, while significant increases with MO-B pretreatment
were observed compared with H2O2 treatment
alone (Fig. 3B).
MO-B ameliorates
H2O2-induced apoptosis in HUVECs
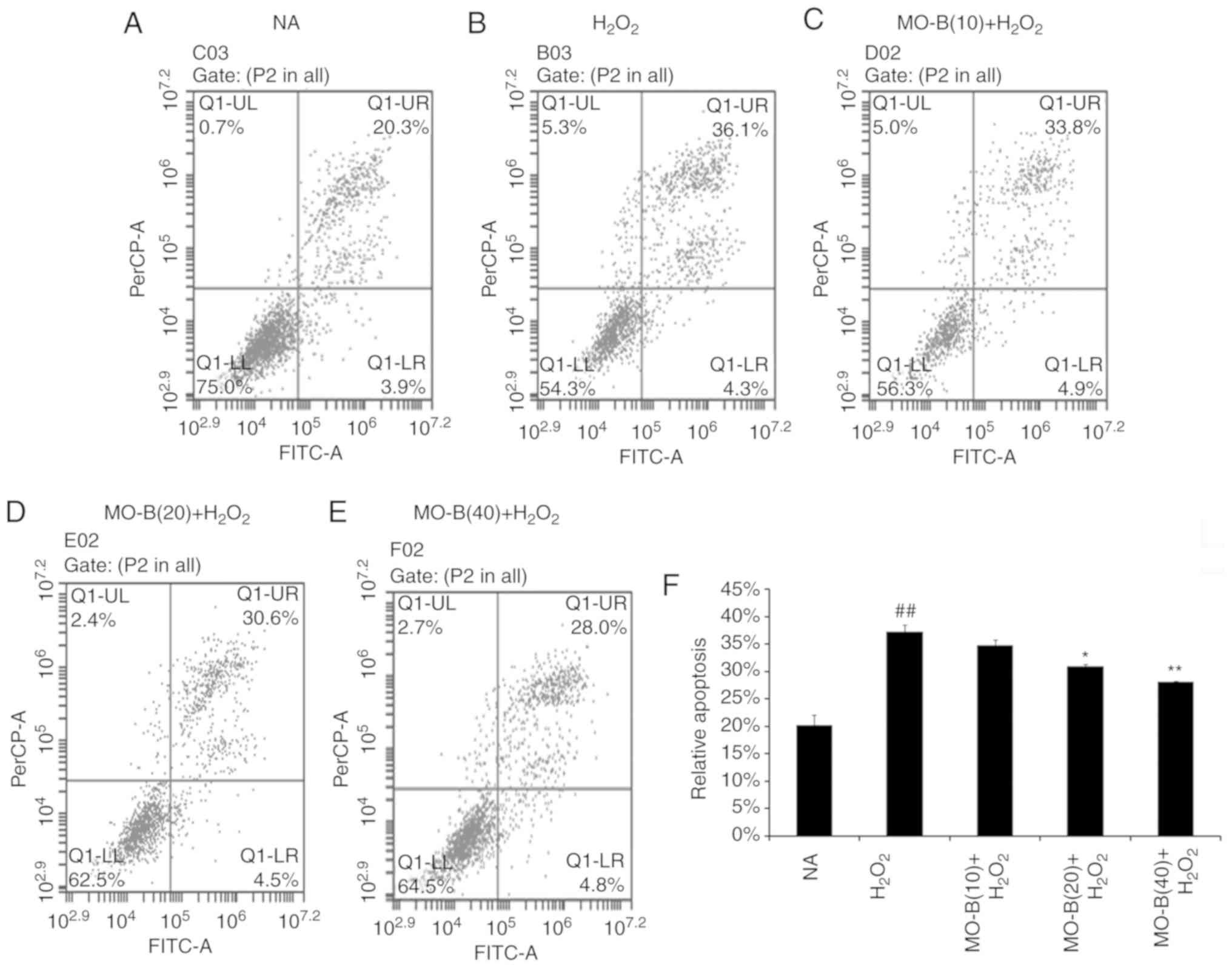
To further illustrate the effects of MO-B on
H2O2-induced vascular endothelial cell
injury, cell apoptosis was measured by the Annexin V-FITC and PI
double-staining method, and the number of apoptotic cells of HUVECs
was determined by flow cytometry. As shown in Fig. 4B, the percentage of double-positive
cells (upper right quadrant; 36.1%) was significantly increased
compared with the normal group (20.3%) (Fig. 4A and F). This level of apoptosis
was significantly ameliorated by the three concentrations of MO-B
tested, ranging from 33.8 to 20.8%, compared with
H2O2 treatment alone (Fig. 4C-F). These results indicate that
MO-B could protect HUVECs from
H2O2-induced apoptosis.
Apoptosis is an orderly cell death process regulated by
apoptosis-associated genes, including the Bcl-2 family and caspases
family and cytochrome c. The Bcl-2 family is composed of
proteins that regulate apoptosis by inducing (pro-apoptotic) or
inhibiting (anti-apoptotic) this process (23). The Bcl-2 family plays a dual role
in the regulation of apoptosis (24). Thus, the present study detected the
mRNA expression of Bcl-2 in HUVECs, which prevents apoptosis, and
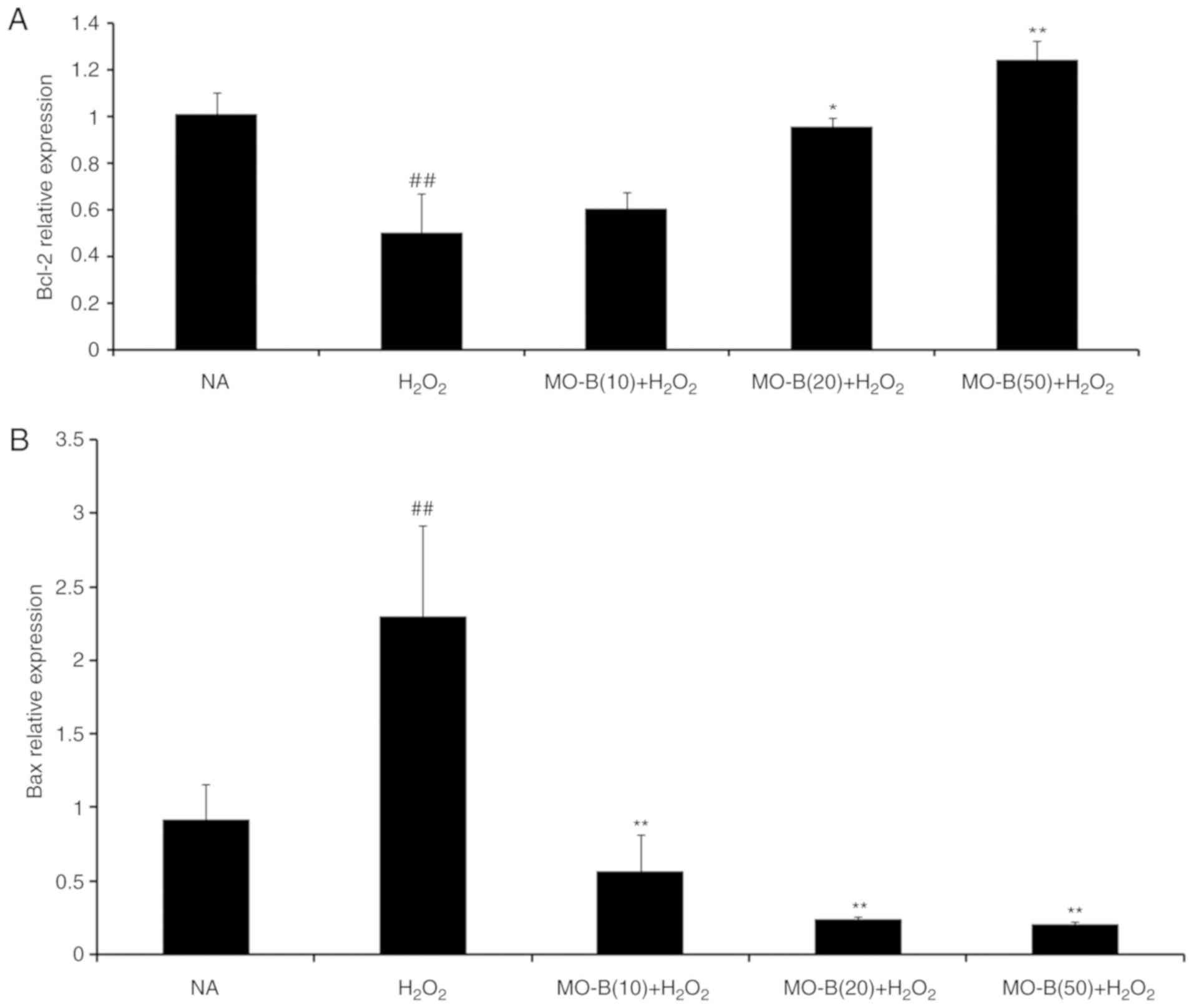
that of Bax, which promotes apoptosis. As presented in Fig. 5A and B, H2O2
significantly promoted Bax mRNA expression, but suppressed that of
Bcl-2 in HUVECs compared with the control group. Pretreatment of
MO-B led to significantly downregulated Bax and increased Bcl-2
expression compared with H2O2 treatment
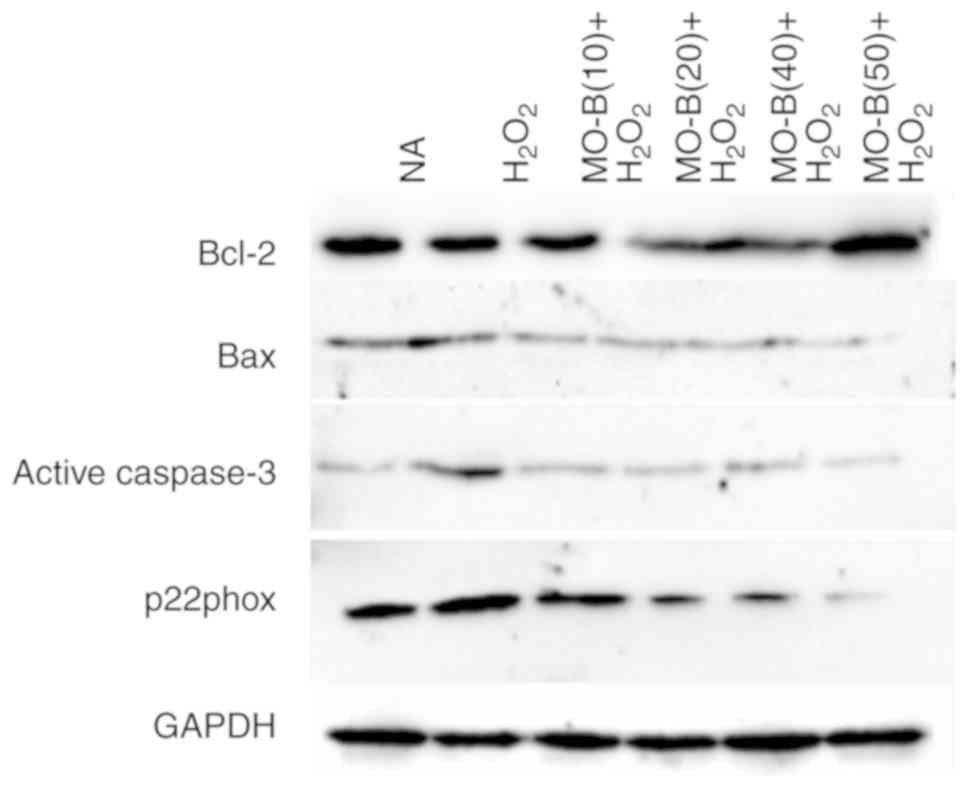
alone. Similar results were observed for protein expression in
which H2O2 alone markedly increased the level
of Bax but decreased the Bcl-2 protein expression, while MO-B could
dose-dependently ameliorate the effects of
H2O2 (Fig.
6). The results indicated that H2O2
induced cell apoptosis, which was reversed with MO-B
pretreatment.
Furthermore, caspases belong to a family of cysteine
proteases that are key mediators of programmed cell death or
apoptosis (25). Caspase-3 is a
marker of apoptosis, and the initiator and executor of this process
(26). I addition, caspase-3 is
the hub of various apoptotic signal transduction pathways outside
the cell, and is the most important apoptotic protease, which is
activated in the early stage of apoptosis (27). Activated caspase-3 causes a series
of cascade reactions, which eventually lead to apoptosis. Hence,
the activity of caspase-3 in HUVECs was examined. In Fig. 5C, H2O2
significantly increased caspase-3 mRNA expression in HUVECs
compared with the control group. Pretreatment with MO-B (10, 20 or
50 µM) significantly inhibited caspase-3 mRNA expression compared
with H2O2 treatment alone. Western blotting
also revealed the same trend in active caspase-3 protein
expression. There was an increase in active caspase-3 expression
caused by H2O2 treatment, which could be
inhibited by administration of MO-B (Fig. 6). Collectively, these results
further suggested the pivotal role of caspase-3 activation in
H2O2-induced apoptosis, as well as the
anti-apoptotic effect of MO-B.
MO-B activates NADPH signaling via
inhibition of p22phox overexpression
NADPH oxidase serves a key role in ROS generation,
and p22phox is a modulatory subunit of NADPH oxidase (28). Thus, the effects of MO-B on
H2O2-induced p22phox mRNA and protein
expression in HUVECs were examined. A notable increase in p22phox
mRNA levels was observed in the H2O2 group.
Pretreatment of HUVECs with MO-B (>10 µM) significantly reversed
H2O2-induced p22phox expression (Fig. 5D). A similar result was also
observed for protein expression. H2O2 alone
markedly increased the level of p22phox protein expression by
2-fold compared with the control group, while MO-B could
dose-dependently attenuate its protein expression (Fig. 6). These results suggested that MO-B
may exert its effects on p22phox expression by modulating the NADPH
pathway.
Discussion
In the present study, 1 mM
H2O2 effectively induced the apoptosis of
HUVECs, while MO-B protected HUVECs from
H2O2-induced apoptosis possibly via the
stimulation of the NADPH oxidase pathway, acting as an
antioxidant.
There are >110 homoisoflavonoid compounds, which
are a particular type of flavonoid, isolated from natural materials
(29). Pharmacological activity
studies have shown that homoisoflavonoids have anti-inflammatory
(30,31), antioxidative (5,32,33),
anti-tumor (34,35), antimicrobial properties (36). Other type of homoisoflavonoids,
such as sappanin-type homoisoflavonoids from the fibrous roots of
Polygonatum odoratum (Mill.) Druce, were reported to be
potent glucose transporter 2 inhibitors with glucose-lowering
properties (37). These compounds
cause notable reductions in cardiovascular events and have been
reported as potential cardiovascular drugs, which have notable
antiangiogenic effects (38,39).
MO-B is a homoisoflavonoid that, besides antioxidant (40) and anti-tumor activity (17), appears to have protective effects
on the cardiovascular system (16). MO-A, a structural analogue of MO-B,
can suppress ischemia/reperfusion-induced myocardial apoptosis in
mice (41), and protects against
cerebral ischemia/reperfusion injury and attenuates blood-brain
barrier disruption in vitro (42). Similarly, MO-B caused the
inhibition of HIF-1α activity (16). As HIF-1α was demonstrated to induce
the production of the pleiotropic proinflammatory cytokine
migration inhibitory factor (MIF) in macrophages (43), and MIF is implicated in several
immunoinflammatory and autoimmune diseases and cancer (44–48),
MO-B may negatively regulate MIF in several immunoinflammatory and
autoimmune diseases. MO- A and MO-B are the major contributors to
the total homoisoflavonoid content in Ophiopogon japonicas
(40). Previous pharmacological
studies have focused their attention on MO-A as a natural drug able
to inhibit the progression of cardiovascular diseases (40,41).
At present, there few studies have investigated MO-B compounds as
of the low abundance in plants (16).
Apoptosis is a complex process that occurs via a
series of physiological activities in cells, including lipid
peroxidation, apoptotic gene and protein expression and apoptotic
body formation. MDA can be applied as a biomarker to evaluate the
degree of lipid peroxidation on the cell membrane (49), which is generated by oxidative
stress in the organism (50). In
the HUVEC oxidative damage model, increased levels of MDA were
linked to greater toxicity and could affect normal cell functions
by interacting with phospholipid proteins, and accumulating inside
the cells (51), while the
activity of SOD was significantly decreased; the antioxidant
function of SOD is involved in preventing injuries to the cell
membrane due to ROS (52). The
percentage of apoptotic cells decreased in a dose-dependent manner
in the presence of MO-B, indicating that MO-B can prevent the
apoptosis of HUVECs and help to maintain cell integrity and
activity. These findings suggested that MO-B protected cells from
apoptosis via intracellular antioxidant enzymes. Furthermore,
apoptotic gene expression was induced, including that of Bcl-2/Bax
and caspase-3. The primary role of Bcl-2 family members is the
regulation of apoptosis, and the ratio of Bcl-2/Bax determines the
fate of cells upon exposure to various stimuli (53). In the present study, the mRNA
expression levels of Bax were downregulated, while those of Bcl-2
were upregulated when cells were exposed to MO-B and
H2O2. These findings were confirmed by
measuring protein expression via western blot analysis. Caspase-3
is an initiator and executioner of apoptosis (54). Our study reported that MO-B
suppress the apoptosis of endothelial cells caused by
H2O2 via downregulation of
cleaved-caspase-3.
The primary catalytic function of NADPH oxidase is
to generate ROS (55). The enzyme
is stimulated by hypertension, hypercholesterolemia, diabetes and
ageing, and once activated, it causes oxidative stress, endothelial
dysfunction and vascular inflammation, which are the early steps of
arterial remodeling and atherogenesis (56). p22phox is an important component of
NADPH oxidase, and can activate this enzyme (57), serving a critical role under
oxidative stress in cardiovascular disease (58). In previous studies, the expression
of p22phox both at the protein and mRNA level was closely followed
by the release of ROS (51), and
p22phox was upregulated under H2O2
stimulation, while antioxidant treatment with vitamin C or
diphenyleneiodonium abrogated thrombin-induced ROS production and
p22phox expression (59). In the
present study, using a cell culture model, it was observed that
MO-B could significantly reduce ROS levels in HUVECs, suggesting
that MO-B may play an antioxidative role by inhibiting NADPH
oxidase activity. Compared with the
H2O2-induced group, the mRNA and protein
expression levels of p22phox in the MO-B group exhibited a
dose-dependent decrease, which supported our hypothesis. This
suggested that MO-B may exert a protective effect via NADPH oxidase
on HUVECs induced by H2O2; however, further
investigation is required.
In conclusion, homoisoflavonoids from Radix
Ophiopogonis may be potential therapeutic agents for treating
cardiovascular disease. Homoisoflavonoids are a particular type of
flavone compound, whose parental structure has an additional carbon
atom than isoflavones. This type of compound is notably rare in
plants, and is mainly distributed among Ophiopogon, Scilla,
Eucomis and Muscari (29). Although the protective effects of
MO-B on damaged endothelial cells have been confirmed in the
present study, the associated pathological mechanisms remain
unknown. In addition, a recent study investigated this compound
in vivo, using a diet with low-dose and long-term
concentrations of MO-B daily for 2 weeks (40). The clinical application of Radix
Ophiopogonis has been reported; however, previous pharmacological
research on Radix Ophiopogonis has mainly focused on its effects,
while the activity of its individual chemical components was rarely
studied (11). The association
between the monomer components and the target is not clear; thus,
further studies on the efficacy of the monomers are of great
importance for the safety, effectiveness, controllability and
stability of clinical application of this drug.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Zhejiang
Provincial Natural Science Foundation of China (grant no.
LY16C020002) and Zhejiang Provincial Science and Technology Project
(grant no. 2018C02042).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and LW made substantial contributions to the
conception and design of the study. LW and YQ performed the
experiments. YW, BL and LW made substantial contributions to the
acquisition, analysis and interpretation of the data and wrote the
paper. MB and RF designed the experiment, analyzed the data, and
contributed reagents, materials and analysis tools. MB, LW and YZ
reviewed and edited the manuscript. All authors read and approved
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Benjamin EJ, Blaha MJ, Chiuve SE, Cushman
M, Das SR, Deo R, de Ferranti SD, Floyd J, Fornage M, Gillespie C,
et al: Heart disease and stroke statistics-2017 update: A report
from the American Heart Association. Circulation. 135:e146–e603.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lockshin RA and Zakeri Z: Programmed cell
death and apoptosis: Origins of the theory. Nat Rev Mol Cell Biol.
2:545–550. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dimmeler S, Hermann C and Zeither AM:
Apoptosis of endothelial cells: Contribution to the pathophysiology
of atherosclerosis. Eur Cytokine Netw. 9:697–698. 1998.PubMed/NCBI
|
|
4
|
Sharifpanah F and Sauer H: Reactive oxygen
species, oxidative stress, and cardiovascular diseasesOxidative
Stress and Antioxidant Protection: The Science of Free Radical
Biology and Disease. Armstrong D and Stratton RD: John Wiley &
Sons Inc.; Hoboken, NJ: pp. 281–306. 2016, View Article : Google Scholar
|
|
5
|
Zhu YZ, Huang SH, Tan BK, Sun J, Whiteman
M and Zhu YC: Antioxidants in Chinese herbal medicines: A
biochemical perspective. Nat Prod Rep. 21:478–489. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu LY, Zheng GQ and Wang Y: An overview of
systematic reviews of shenmai injection for healthcare. Evid Based
Complement Alternat Med. 2014:8406502014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen HD, Xie YM, Wang LX and Wu JB:
Systematic review of efficacy and safety of shenmai injection for
chronic heart failure. Zhongguo Zhong Yao Za Zhi. 39:3650–3661.
2014.(In Chinese). PubMed/NCBI
|
|
8
|
Jun F and Xu Z: Advancement in research of
pharmacological functions of Radix Ophiopogonis on cardiovascular
system. J Nanjing Univ Tradit Chin Med. 22:270–272. 2006.
|
|
9
|
Zhao M, Xu W, Shen HY, Shen PQ, Zhang J,
Wang DD, Xu H, Wang H, Yan TT, Wang L, et al: Comparison of
bioactive components and pharmacological activities of
Ophiopogon japonicas extracts from different geographical
origins. J Pharm Biomed Anal. 138:134–141. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu BY: Exploration on the modern research
methodology of traditional chinese medicine, basing on the systemic
research of Radix Ophiopogonis. Chin J Nat Med Jan. 5:10–14.
2007.
|
|
11
|
Chen MH, Chen XJ, Wang M, Lin LG and Wang
YT: Ophiopogon japonicas-A phytochemical, ethnomedicinal and
pharmacological review. J Ethnopharmacol. 181:193–213. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fan XH, Wang Y and Cheng YY: LC/MS
fingerprinting of Shenmai injection: A novel approach to quality
control of herbal medicines. J Pharm Biomed Anal. 40:591–597. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gu SL, Xu SS, Ji K, Yang QH, Lu WW, Jia YS
and Wang N: Effects of maidong on experimental myocardial
infarction and submicrostructure in myocardial hypoxia. Shanghai J
Tradit Chin Med. 3:44–45. 1983.
|
|
14
|
Wang Y, Liu F, Liang Z, Peng L, Wang B, Yu
J, Su Y and Ma C: Homoisoflavonoids and the antioxidant activity of
Ophiopogon japonicus root. Iran J Pharm Res. 16:357–365.
2017.PubMed/NCBI
|
|
15
|
Ito Y, Kanamaru AA and Akihiro T: A novel
agent, methylophiopogonanone B, promotes Rho activation and tubulin
depolymerization. Mol Cell Biochem. 297:121–129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fujii M, Egawa K, Hirai Y, Kondo M, Fujii
K, Uekusa H, Akita H, Nose K, Toriizuka K and Ida Y:
Dihydrochalcone designed from methylophiopogonanone B strongly
inhibits hypoxia-inducible factor (HIF)-1α activity. Heterocycles.
78:2061–2065. 2009. View Article : Google Scholar
|
|
17
|
Ito Y, Kanamaru A and Tada A: Effects of
methylophiopogonanone B on melanosome transfer and dendrite
retraction. J Dermatol Sci. 42:68–70. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang KW, Zhang H, Shen LQ and Wang W:
Novel steroidal saponins from liriope graminifolia (Linn.)
baker with anti-tumor activities. Carbohydr Res. 346:253–258. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou Y, Wang L, Liu T, Mao Z, Ge Q and Mao
J: Isolation of homoisoflavonoids from the fibrous roots of
Ophiopogon japonicus by recycling high-speed
counter-currentchromatography and online antioxidant activity
assay. Acta Chromatogr. Oct 14–2018.(Epub ahead of print).
doi.org/10.1556/1326.2018.00509. View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gaweł S, Wardas M, Niedworok E and Wardas
P: Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek.
57:453–455. 2004.(In Polish). PubMed/NCBI
|
|
22
|
Zelko IN, Mariani TJ and Folz RJ:
Superoxide dismutase multigene family: A comparison of the CuZn-SOD
(SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures,
evolution, and expression. Free Radic Biol Med. 33:337–349. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cleary ML, Smith SD and Sklar AJ: Cloning
and structural analysis of cDNAs for bcl-2 and a hybrid
bcl-2/immunoglobulin transcript resulting from the t(14;18)
translocation. Cell. 47:19–28. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gross A, Mcdonnell JM and Korsmeyer SJ:
Bcl-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Enari M, Sakahira H, Yokoyama H, Okawa K,
Iwamatsu A and Nagata S: A caspase-activated DNase that degrades
DNA during apoptosis, and its inhibitor ICAD. Nature. 391:43–50.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia F, Wang C, Jin Y, Liu Q, Meng Q, Liu K
and Sun H: Luteolin protects HUVECs from TNF-α-induced oxidative
stress and inflammation via its effects on the Nox4/ROS-NF-κB and
MAPK pathways. J Atheroscler Thromb. 21:768–783. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang HB, Huang J, Guo MJ, Zou P and Tian
XQ: Recent advances in the study of natural homoisoflavonoids. Yao
Xue Xue Bao. 42:118–126. 2007.(In Chinese). PubMed/NCBI
|
|
30
|
Hung TM, Thu CV, Dat NT, Ryoo SW, Lee JH,
Kim JC, Na M, Jung HJ, Bae K and Min BS: Homoisoflavonoid
derivatives from the roots of Ophiopogon japonicus and their
in vitro anti-inflammation activity. Bioorg Med Chem Lett.
20:2412–2416. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Damodar K, Lee J, Kim JK and Jun JG:
Synthesis and in vitro evaluation of homoisoflavonoids as potent
inhibitors of nitric oxide production in RAW-264.7 cells. Bioorg
Med Chem Lett. 28:2098–2102. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siddaiah V, Maheswara M, Venkata Rao C,
Venkateswarlu S and Subbaraju GV: Synthesis, structural revision,
and antioxidant activities of antimutagenic homoisoflavonoids from
Hoffmanosseggia intricata. Bioorg Med Chem Lett. 17:1288–1290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou YF, Qi J, Zhu DN and Yu BY:
Homoisoflavonoids from Ophiopogon japonicus and its oxygen
free radicals (OFRs) scavenging effects. Chin J Nat Med. 6:201–204.
2008. View Article : Google Scholar
|
|
34
|
El-Elimat T, Rivera-Chávez J, Burdette JE,
Czarnecki A, Alhawarri MB, Al-Gharaibeh M, Alali F and Oberlies NH:
Cytotoxic homoisoflavonoids from the bulbs of Bellevalia flexuosa.
Fitoterapia. 127:201–206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Duan CL, Kang ZY, Lin CR, Jiang Y, Liu JX
and Tu PF: Two new homoisoflavonoids from the fibrous roots of
Ophiopogon japonicus (Thunb.) Ker-Gawl. J Asian Nat Prod
Res. 11:876–879. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alali F, El-Elimat T, Albataineh H,
Al-Balas Q, Al-Gharaibeh M, Falkinham JO III, Chen WL, Swanson SM
and Oberlies NH: Cytotoxic homoisoflavones from the bulbs of
Bellevalia eigii. J Nat Prod. 78:1708–1715. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang H, Fowler MI, Messenge DJ, Terry LA,
Gu X, Zhou L, Liu R, Su J, Shi S, Ordaz-Ortiz JJ, et al:
Homoisoflavonoids are potent glucose transporter 2(GLUT2)
inhibitors: A potential mechanism for the glucose-lowering
properties of Polygonatum odoratum. J Agric Food Chem.
66:3137–3145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee B, Sun W, Lee H, Basavarajappa H,
Sulaiman RS, Sishtla K, Fei X, Corson TW and Seo SY: Design,
synthesis and biological evaluation of photoaffinity probes of
antiangiogenic homoisoflavonoids. Bioorg Med Chem Lett.
26:4277–4281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Amin SA, Adhikari N, Gayen S and Jha T:
Homoisoflavonoids as potential antiangiogenic agents for retinal
neovascularization. Biomed Pharmacother. 95:818–827. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He F, Xu BL, Chen C, Jia HJ, Wu JX, Wang
XC, Sheng JL, Huang L and Cheng J: Methylophiopogonanone A
suppresses ischemia/reperfusion-induced myocardial apoptosis in
mice via activating PI3K/Akt/eNOS signaling pathway. Acta Pharmacol
Sin. 37:763–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lin M, Sun W, Gong W, Zhou Z, Ding Y and
Hou Q: Methylophiopogonanone a protects against cerebral
ischemia/reperfusion injury and attenuates blood-brain barrier
disruption in vitro. PLoS One. 10:e01245582015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Richard V, Kindt N and Saussez S:
Macrophage migration inhibitory factor involvement in breast cancer
(Review). Int J Oncol. 47:1627–1633. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Günther S, Fagone P, Jalce G, Atanasov AG,
Guignabert C and Nicoletti F: Role of MIF and D-DT in
immune-inflammatory, autoimmune, and chronic respiratory diseases:
From pathogenic factors to therapeutic targets. Drug Discov Today.
24:428–439. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fagone P, Mazzon E, Cavalli E, Bramanti A,
Petralia MC, Mangano K, Al-Abed Y, Bramati P and Nicoletti F:
Contribution of the macrophage migration inhibitory factor
superfamily of cytokines in the pathogenesis of preclinical and
human multiple sclerosis: In silico and in vivo evidences. J
Neuroimmunol. 322:46–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Presti M, Mazzon E, Basile MS, Petralia
MC, Bramant A, Colletti G, Bramanti P, Nicoletti F and Fagone P:
Overexpression of macrophage migration inhibitory factor and
functionally-related genes, D-DT, CD74, CD44, CXCR2 and CXCR4, in
glioblastoma. Oncol Lett. 16:2881–2886. 2018.PubMed/NCBI
|
|
46
|
Mangano K, Mazzon E, Basile MS, Marco R,
Bramanti P, Mammana S, Petralia MC, Fagone P and Nicoletti F:
Pathogenic role for macrophage migration inhibitory factor in
glioblastoma and its targeting with specific inhibitors as novel
tailored therapeutic approach. Oncotarget. 9:17951–17970. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kindt N, Journe F, Laurent G and Saussez
S: Involvement of macrophage migration inhibitory factor in cancer
and novel therapeutic targets. Oncol Lett. 12:2247–2253. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lin Y, Zhu D, Qi J, Qin M and Yu B:
Characterization of homoisoflavonoids in different cultivation
regions of Ophiopogon japonicus and related antioxidant
activity. J Pharm Biomed Anal. 52:757–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yapislar H and Taskin E: L-carnosine
alters some hemorheologic and lipid peroxidation parameters in
nephrectomized rats. Med Sci Monit. 20:399–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tsikas D: Assessment of lipid peroxidation
by measuring malondialdehyde (MDA) and relatives in biological
samples: Analytical and biological challenges. Anal Biochem.
524:13–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jin Y, Liu K, Peng J, Wang C, Kang L,
Chang N and Sun H: Rhizoma Dioscoreae Nipponicae polysaccharides
protect HUVECs from H2O2-induced injury by
regulating PPARγ factor and the NADPH oxidase/ROS-NF-κB signal
pathway. Toxicol Lett. 232:149–158. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Atig F, Raffa M, Ali HB, Abdelhamid K,
Saad A and Ajina M: Altered antioxidant status and increased lipid
per-oxidation in seminal plasma of tunisian infertile men. Int J
Biol Sci. 8:139–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Boatright KM and Salvesen GS: Mechanisms
of caspase activation. Curr Opin Cell Biol. 15:725–731. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zafari AM, Ushio-Fukai M, Akers M, Yin Q,
Shah A, Harrison DG, Taylor WR and Griendling KK: Role of
NADH/NADPH oxidase-derived H2O2 in angiotensin II-induced vascular
hypertrophy. Hypertension. 32:488–495. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Drummond GR, Selemidis S, Griendling KK
and Sobey CG: Combating oxidative stress in vascular disease: NADPH
oxidases as therapeutic targets. Nat Rev Drug Discov. 10:453–471.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Schramm A, Matusik P, Osmenda G and Guzik
TJ: Targeting NADPH oxidases in vascular pharmacology. Vascul
Pharmacol. 56:216–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
San José G, Fortuño A, Beloqui O, Díez J
and Zalba G: NADPH oxidase CYBA polymorphisms, oxidative stress and
cardiovascular diseases. Clin Sci (Lond). 114:173–182. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Djordjevic T, Pogrebniak A, BelAiba RS,
Bonello S, Wotzlaw C, Acker H, Hess J and Görlach A: The expression
of the NADPH oxidase subunit p22phox is regulated by a
redox-sensitive pathway in endothelial cells. Free Radic Biol Med.
38:616–630. 2005. View Article : Google Scholar : PubMed/NCBI
|