Introduction
Osteoarthritis (OA) is identified as the degradation
of articular cartilage, and it affects >43 million people
globally. It is the most common cause of disability and
disproportionately affects the elderly (1,2). OA
is associated with a number of risk factors, including ageing,
obesity and acetabular dysplasia, and has a hereditary aspect
(3,4). Deterioration of the cartilage is
generally accepted as the primary cause of OA occurrence.
Chondrocytes are considered to be the target of biomechanical
factors leading to abnormal functional changes in these cells,
resulting in physiological changes that cause OA (5). OA is commonly treated with joint
replacement surgery and anti-inflammatory drugs, although the
inflammatory drugs have several side effects (6). Improving the understanding of the
underlying molecular mechanisms of inflammatory injury of cartilage
cells in OA may result in the development of improved therapeutic
options.
Long noncoding RNAs (lncRNAs) are noncoding
transcripts that are >200 nucleotides in length and lack protein
encoding capacity (7). Although
lncRNAs were initially thought to be transcriptional noise, they
are now regarded as molecules with diverse functional roles in
regulating a range of cellular functions. Different lncRNAs have
exhibited differential expression patterns in different diseases
and are involved in various developmental and pathological
conditions of a number of diseases (8). A recent study suggested that lncRNAs
serve a vital role in OA pathogenesis (9). lncRNA NR024118 is a recently
identified lncRNA that is closely associated with the
pathophysiology of a number of diseases, including cancer,
cardiovascular diseases, neural diseases and rheumatoid arthritis
(RA). However, the potential effects of lncRNA NR024118 in OA
remain unknown.
NF-κB is a well-studied and key inflammatory
molecule. Activation of the NF-κB pathway results in the production
of pro-inflammatory cytokines, including tumor necrosis factor α,
interleukin (IL)-1β and IL-6 (10). Nuclear factor erythroid-2 related
factor 2 (Nrf2) is an important transcription factor and master
regulator of the cellular response to oxidative stress and the
inflammatory process, which increases the expression of antioxidant
and cytoprotective enzymes and downstream proteins including
catalase (CAT), superoxide dismutase (SOD), quinone
oxidoreductase-1 (NQO1) and heme oxygenase-1 (HO-1) (11), and the Nrf2 pathway serves an
important role in lipopolysaccharide (LPS)-induced inflammatory
injury (12).
In the present study, the potential effects of
NR024118 on apoptosis and inflammatory damage in LPS-treated
chondrocytes were measured. Furthermore, the effects of NR024118 on
LPS-induced activation of the NF-κB signaling pathways and
inhibition of the Nrf2 signaling pathway were examined. The results
of the present study may provide insight into the identification of
novel therapeutic and diagnostic targets for diagnosing and
treating patients with OA.
Materials and methods
Cell culture and treatment
The murine chondrogenic ATDC5 cell line was obtained
from the Cell Bank Cell Engineering Division, RIKEN BioResource
Research Center and cultured in DMEM/Ham Nutrient Mixture F12
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified incubator with 5% CO2. Cells were treated
with 5 µg/ml LPS (5 mg/ml; Sigma-Aldrich; Merck KGaA) for 5 h to
stimulate inflammatory injury.
Transfection
The NR024118 sequence
(TCAAAGACCACCACCATCTTCCTCAATGGCAACCGCGAGCGGCCCTTGGATGTGTTTTGTGACATGCAGACTGACGGAGGAGGTTGGCTGGTGTTCCAGCGCCGCATGGACGGACAGACAGACTTCTGGAGAGACTGGGAGGAGTACGCCCATGGTTTTGGGAACATCTCCAGGGAATTCTGGCTGGGCAATGAGGCCCTTCACAGCCTCACGCAGGCTGGAGACTACTCTATGCGTGTGGACCTGCGGGCCGGAAAGGAAGCCGTGTTCGCCCAGTATGACTTCTTCCGAGTAGACTCAGCGAAGGAGAACTATCGTCTACACCTAGGGGGCTACCATGGGACCGCGGGTGACTCTATGAGCTACCACAGCGGCAGTGCCTTTTCTGCCCGTGATCGAGACCCCAATAACTTGCTCATCTCCTGCGCTGTCTCCTATCGTGGGGCTTGGTGGTACAGGGACTGTCACTACGCCAATCTCAATGGGCTCTATGGGAGCACAGTGGATCACCAGGGAGTGAGCTGGTACCACTGGAAGGGCTTCGAGTTCTCGGTGCCCTTCACGGAAATGAAGCTGAGACCCAGAAACTTCCAGGCCCCCACCAGGGGCACCTGAGCCTGCTGCCCACCTCACTCACACCCTGGTATGACTGCCGAGCACTGAGGGGTTGTGCCCAGAGAAGAGCCAGTGTGTCTCTACTGTGCCTAGCTCACCGAGGAAGCCTTCTCTGCCACAGTCTCACAGCACCATGTTTACAGGGGGGAGGGGAGGGAAATGGAGCAATAAAGGAGAA)
was synthesized and subcloned into a pcDNA3.1 vector (Invitrogen;
Thermo Fisher Scientific, Inc.). The pcDNA3.1-NR024118 (NR024118)
or empty vector was transfected into chondrocytes using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols, and
the concentration is as followed: pcDNA 3.1: Lipofectamine=2 µg: 4
µl. After 48 h, reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) was used to evaluate the efficiency of the
transfections.
Cell viability assay
Cell viability assay was assessed using a Cell
Counting Kit-8 (CCK-8) assay Dojindo Molecular Technologies, Inc.).
Briefly, cells (5×103 cells/well) were seeded in a
96-well cell culture plate following LPS treatment and/or
transfection with pcDNA3.1-NR024118 and 20 µl CCK-8 solution was
added into each well and incubated at 37°C for 2 h. The absorbance
at 450 nm of each well was measured using a microplate reader
(Bio-Rad Laboratories, Inc.).
Cell apoptosis assay
Cell apoptosis was detected using fluorescein
isothiocyanate (FITC)-Annexin V/propidium iodide (PI) staining (BD
Biosciences). Briefly, following LPS treatment and/or transfection
with pcDNA3.1-NR024118, ATDC5 cells were collected and stained with
10 µl Annexin V-FITC and 5 µl PI in the dark at 37°C for 25 min.
Subsequently, the percentage of apoptotic cells were determined
using flow cytometry (Guava Technologies, Inc.; Luminex
Corporation). FlowJo software version 10.0 (FlowJo LLC) was used to
analyze the data.
Cytokine analysis
The concentrations of IL-1β, IL-6 and IL-18 were
determined using specific ELISA kits (Mouse IL-1β/ IL-1F2
Quantikine ELISA kit, cat. no. MLB00C; Mouse IL-6 Quantikine ELISA
kit, cat. no. M6000B, and Mouse IL-18 ELISA, cat. no. 7625,
respectively; R&D Systems, Inc.,) according to the
manufacturer's protocols.
Measurement of intracellular reactive
oxygen species (ROS)
The intracellular ROS levels were measured using the
probe CM-H2DCFDA (Invitrogen; Thermo Fisher Scientific,
Inc.). Briefly, following LPS treatment and/or transfection with
pcDNA3.1-NR024118, the cells were incubated with DCF-DA at 37°C for
30 min. Subsequently, the cells were trypsinized and suspended in
PBS. Cells were added with PI (1 µg/ml) on ice in the dark, and the
fluorescence intensities were measured at 485 and 535 nm using Cell
Lab Quanta SC MPL flow cytometer (Beckman Coulter, Inc.).
Acquisition and analysis of the data were performed the software of
the flow cytometer.
Measurement of antioxidant enzyme
activity
The activity of SOD was determined using a SOD assay
kit (cat. no. A001-3-2; Nanjing Jiancheng Bioengineering
Institute). MDA levels were determined using an MDA assay kit (cat.
no. A003-1-2; Nanjing Jiancheng Bioengineering Institute) and CAT
levels were examined using a CAT assay kit (cat. no. A007-1-1;
Nanjing Jiancheng Bioengineering Institute) according to the
manufacturers' protocols.
Western blot analysis
ATDC5 cells were lysed using RIPA lysis buffer
(Beyotime Institute of Biotechnology) and supplemented with
protease inhibitors (Roche Diagnostics). The protein concentration
was quantified using a bicinchoninic acid assay kit (Pierce; Thermo
Fisher Scientific, Inc.). A total of 20 µg protein per lane was
resolved using 10% SDS-PAGE and transferred to a polyvinylidene
fluoride membrane (EMD Millipore). The membrane was blocked in 5%
skimmed milk at room temperature for 1 h and incubated with
appropriate primary antibodies (dilution 1:1,000) at 4°C overnight.
The primary antibodies used in the present study were:
Anti-phosphorylated (p)-NF-κB inhibitor β (IκBβ; cat. no. ab59195;
Abcam), anti-IκBβ (cat. no. ab32135; Abcam), anti-NF-κB
transcription factor p65 (p65) antibody (cat. no. 8242; Cell
Signaling Technology, Inc), anti-p-NF-κB p65 antibody (cat. no.
3033; Cell Signaling Technology, Inc), Bcl-2 (cat. no. ab196495;
Abcam), Bax (cat. no. ab182733; Abcam), Bcl-2-like protein 11 (Bim;
cat. no. ab7888; Abcam), Nrf2 (cat. no. 12721; Cell Signaling
Technology, Inc), HO-1 (cat. no. 82206; Cell Signaling Technology,
Inc), NQO1 (cat. no. 3187; Cell Signaling Technology, Inc) and
GAPDH (cat. no. ab181603, Abcam). Subsequently, the membranes were
incubated with horseradish peroxidase-conjugated goat anti-mouse
IgG (dilution 1:2,000; cat. no. sc-2005; Santa Cruz Biotechnology,
Inc.) for 1 h at room temperature. The signals were visualized by
enhanced chemiluminescence kit (GE Healthcare) and Image Lab™
Software version 4.1 (Bio-Rad Laboratories, Inc.) and protein
expression levels were quantified using ImageJ version 1.46
(National Institutes of Health).
RT-qPCR
Total RNA from cultured cells was extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. A total of 2 µg RNA was
reversed transcribed into cDNA using the PrimeScript™ RT reagent
kit with DNA Eraser (Takara Biotechnology Co., Ltd.) according to
the manufacturer's protocol using the following conditions: Initial
incubation at 37°C for 15 min, followed by incubation at 85°C for 5
sec. NR024118 expression was quantified using SYBR1 Green RealTime
PCR Master mix (Invitrogen; Thermo Fisher Scientific, Inc.) in an
ABI PRISM® 7300 PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). PCR reactions were performed using the
following conditions: Initial denaturation at 95°C for 10 min,
followed by 40 cycles of denaturation at 95°C for 15 sec and
anneal/extend at 63°C for 60 sec. The primers sequences were:
NR024118 forward, 5′-GCTGCCCACCTCACTCAC-3′; NR024118 reverse,
5′-CTTTATTGCTCCATTTCCCTC-3′; Nrf2 forward,
5′-TTCCTCTGCTGCCATTAGTCAGTC-3′; Nrf2 reverse,
5′-GCTCTTCCATTTCCGAGTCACTG-3′; NQO-1 forward,
5′-AAGAGCCCTGATTGTACTGGC-3′; NQO-1 reverse,
5′-GCGTCCTTCCTTATATGCTAGAGA-3′; HO-1 forward,
5′-GTGACAGAAGAGGCTAAGACCG-3′; HO-1 reverse,
5′-ACAGGAAGCTGAGAGTGAGGAC-3′; GAPDH forward,
5′-GGGAAACTGTGGCGTGAT-3′; GAPDH reverse, 5′-GAGTGGGTGTCGCTGTTGA-3′.
GAPDH was used as the endogenous control and the data was
quantified using the 2−ΔΔCq method (13).
Statistical analysis
Data are presented as the mean ± standard deviation
from at least 3 experimental repeats. Statistical analysis was
performed using GraphPad Prism v.5 (GraphPad Software, Inc.). A
one-way analysis of variance followed by Tukey's post hoc test or
an unpaired two-tailed t-test was used to analyze the data.
P<0.05 was considered to indicate a statistically significant
difference.
Results
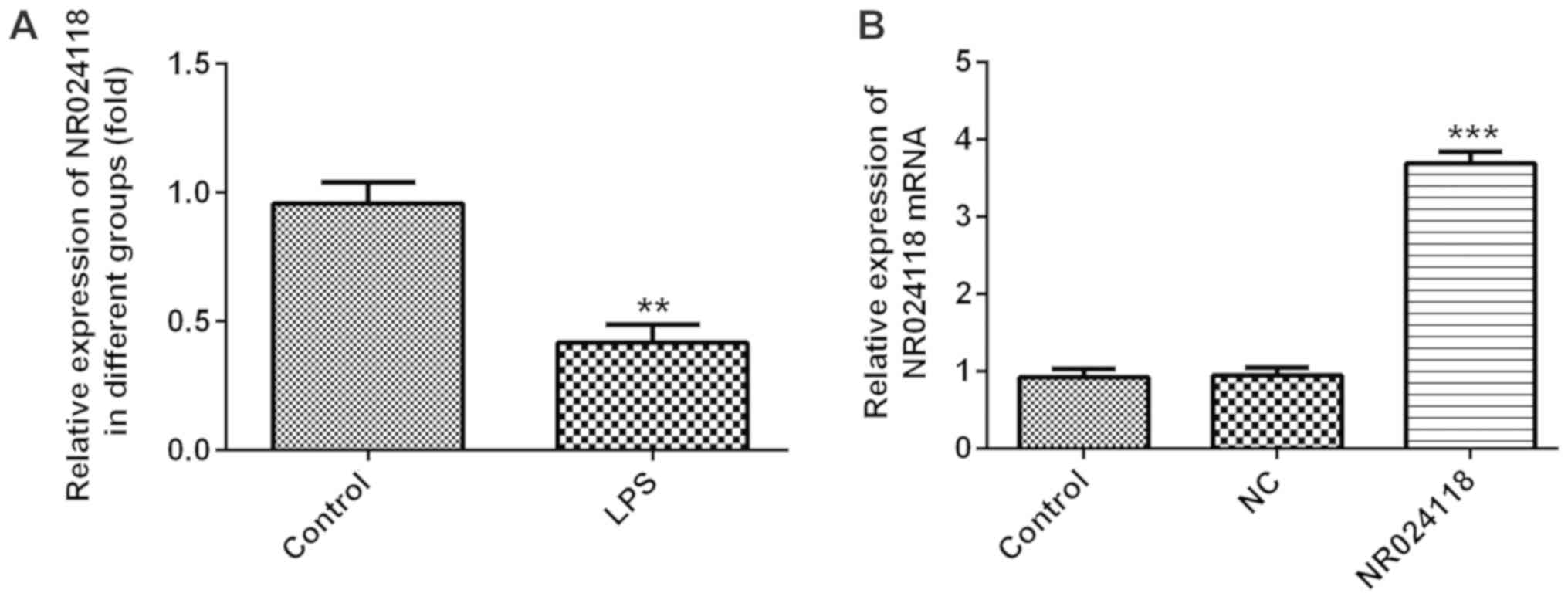
NR024118 expression is downregulated
in LPS-induced in ATDC5 cells
Compared with the control group, NR024118 expression
was significantly decreased in ATDC5 cells following treatment with
LPS (Fig. 1A). To additionally
investigate the biological role of NR024118 in OA, the expression
of NR024118 was overexpressed by transfecting cells with
pcDNA3.1-NR02411. Relative expression of NR024118 was significantly
increased in cells transfected with pcDNA-NR02411 compared with the
control.
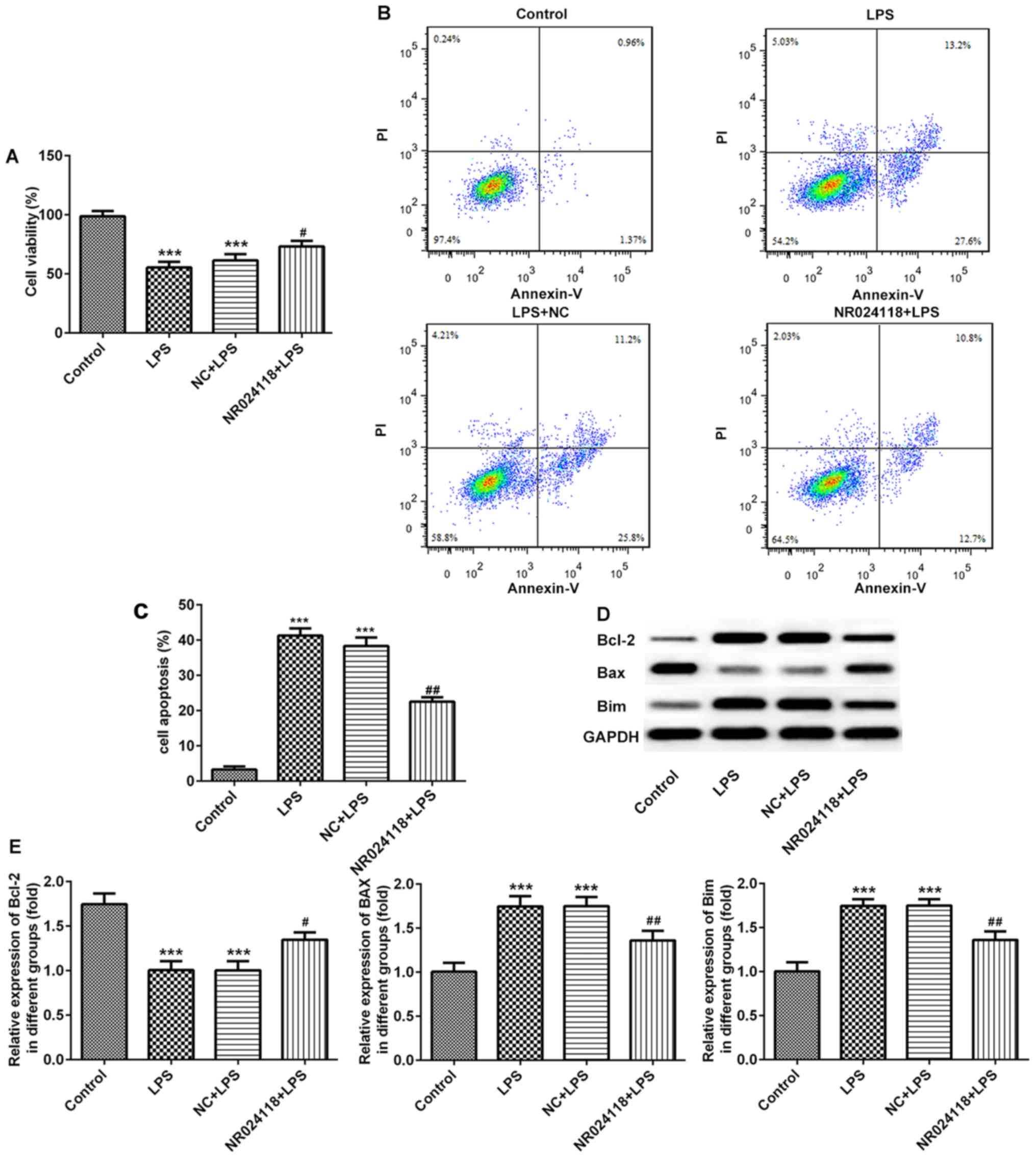
Overexpression of NR024118 attenuates
LPS-induced cell apoptosis in ATDC5 cells
The effect of NR024118 on LPS-induced chondrocyte
apoptosis was determined using a CCK-8 assay and flow cytometry.
The results indicated that the levels of the LPS-induced decrease
in cell viability and induction of apoptosis were decreased in
cells transfected with NR024118 (Fig.
2A-C). In addition, in the LPS-induced cells, Bcl-2 and Bim
expression were upregulated, and Bax was downregulated compared
with the control; these results were reversed in cells transfected
with NR024118.
Overexpression of NR024118 attenuates
LPS-induced inflammatory injury in ATDC5 cells
Following stimulation of cells with 5 µg/ml LPS, the
levels of IL-18, IL-1β and IL-6 were significantly increased
compared with the control group. Overexpression of NR024118
significantly decreased the expression of IL-1β, IL-6 and IL-18 in
LPS-induced cells (Fig. 3A-C). In
addition, ROS production was decreased by NR024118 overexpression
(Fig. 3D). LPS stimulation
significantly increased the levels of MDA (Fig. 3E) and decreased the levels of SOD
(Fig. 3F) and CAT (Fig. 3G) compared with the control group.
These alterations in production and activity were significantly
reversed by the overexpression of NR024118.
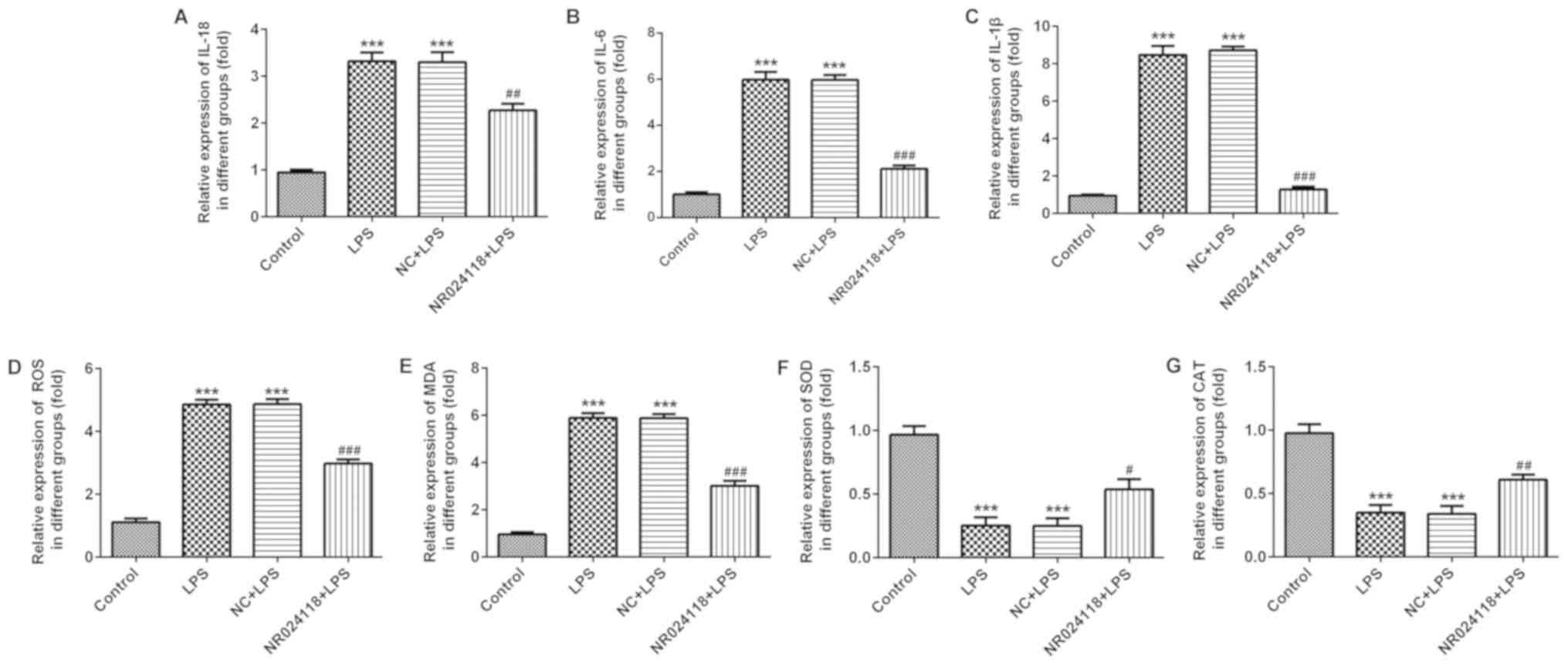 | Figure 3.Overexpression of NR024118 alleviates
LPS-induced inflammatory injury in ATDC5 cells. The concentration
of inflammatory cytokines (A) IL-18, (B) IL-6 and (C) IL-1β in the
culture supernatant of ATDC5 cells was detected by ELISA. (D) ROS,
(E) MDA, (F) SOD and (G) CAT expression were detected by specific
commercial kits. Data are presented as the mean ± standard
deviation. ***P<0.001 vs. control group. #P<0.05,
##P<0.01 and ###P<0.001 vs. LPS group.
LPS, lipopolysaccharide; IL, interleukin; ROS, reactive oxygen
species; SOD, superoxide dismutase; MDA, malondialdehyde; CAT,
catalase; NR024118, pCDNA3.1-NR024118; NC, negative control. |
Overexpression of NR024118 inhibits
LPS-induced NF-κB pathway activation and promotes Nrf2 pathway
activation in ATDC5 cells
Western blot analysis was used to detect the
activation of the NF-κB and Nrf2 signaling pathways in ATDC5 cells
following LPS treatment and/or transfection with pcDNA3.1-NR024118
transfection. As indicated in Fig.
4A-C, LPS treatment resulted in the activation of the NF-κB
pathway by increasing the expression levels of total and p-IκBβ and
total and p-p65. Overexpression of NR024118 significantly inhibited
the LPS-induced NF-κB pathway activation as the expression levels
of total and p-IκBβ and total and p-p65 in ATDC5 cells were
decreased. In addition, LPS treatment significantly activated the
Nrf2 pathway by increasing the protein (Fig. 4D-G) and mRNA (Fig. 4H-J) expression levels of the Nrf2
signaling pathway associated molecules, Nrf2, HO-1 and NQO1.
Overexpression of NR024118 significantly increased the protein and
mRNA expression levels of Nrf2, HO-1 and NQO1.
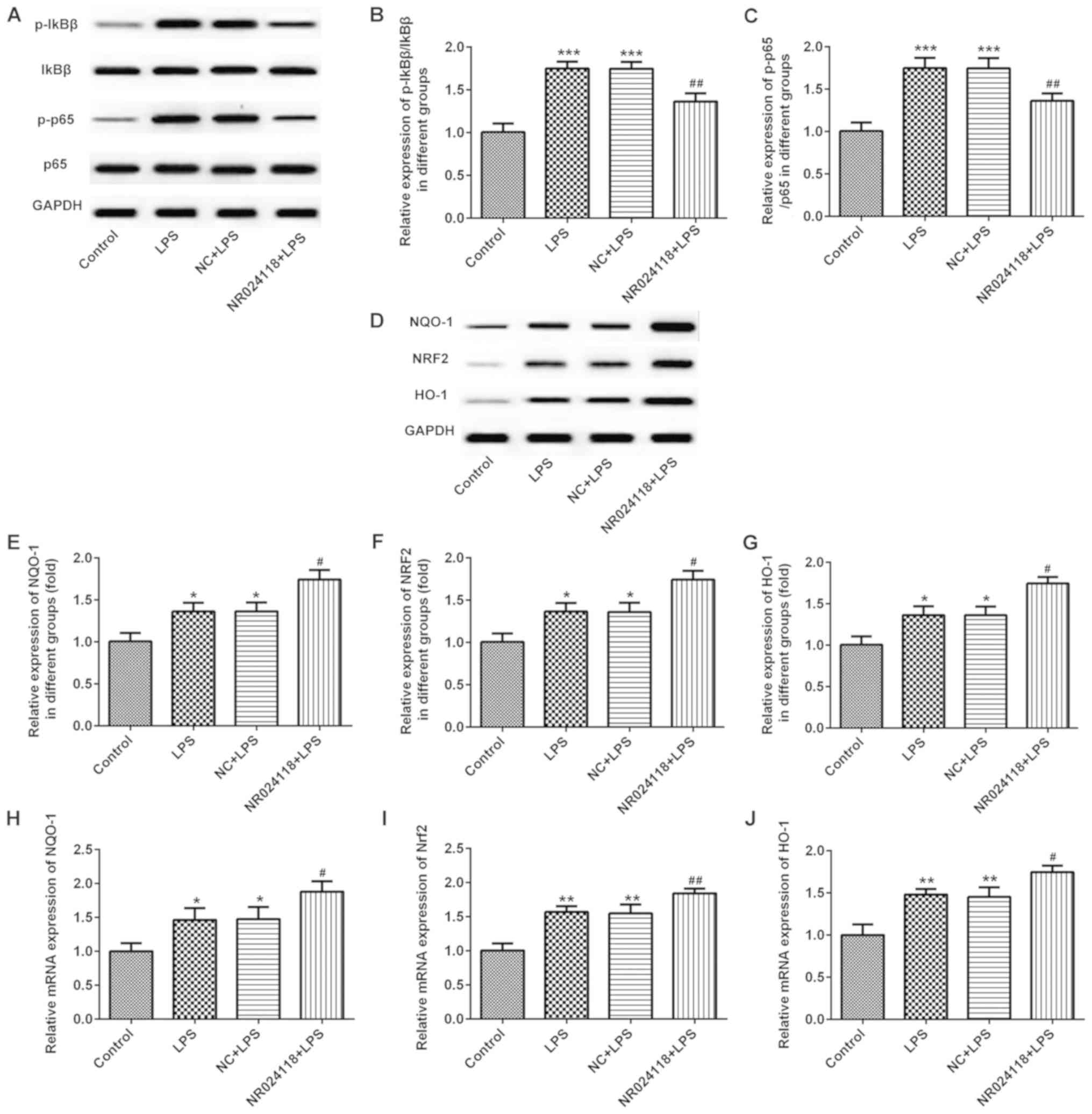 | Figure 4.Overexpression of NR024118 inhibits
LPS-induced NF-κB pathway activation and promotes Nrf2 pathway
activation in ATDC5 cells. (A) Western blot analysis was performed
to evaluate the protein expression levels of p-IκBβ, t-IκBβ, p-p65
and t-p65 following treatment with 5 µg/ml LPS and/or
pCDNA3.1-NR024118 transfection in ATDC5 cells. (B) Quantitative
analyses of the ratios of p/t-IκBβ and (C) p/t-p65 were performed.
(D) Western blot analysis was performed to evaluate the protein
expression of NQO-1, NRF2 and HO-1 following treatment with 5 µg/ml
LPS treatment and/or pCDNA3.1-NR024118 transfection in ATDC5 cells.
Quantitative analyses of the protein expression of (E) NQO-1, (F)
NRF2 and (G) HO-1 were performed. RT-qPCR was conducted to analyze
the mRNA expressions of (H) NQO-1, (I) NRF2 and (J) HO-1 following
treatment with 5 µg/ml LPS and/or pCDNA3.1-NR024118 transfection in
ATDC5 cells. Data are presented as the mean ± standard deviation.
*P<0.05, **P<0.01 and ***P<0.001 vs. control group.
#P<0.05 and ##P<0.01 vs. LPS group.
LPS, lipopolysaccharide; NR024118, pCDNA3.1-NR024118; NC, negative
control; IκBβ, NF-κB inhibitor β; p65, transcription factor p65;
p-, phosphorylated; t-, total; NQO-1, quinone oxidoreductase-1;
NRF2, nuclear factor erythroid-2 related factor 2; HO-1, heme
oxygenase-1. |
Discussion
OA is a widespread joint disorder that plagues
millions of people globally and is a major cause of disability in
the elderly (14). In the present
study, NR024118 expression was significantly increased in
LPS-induced ATDC5 cells. Overexpression of NR024118 reversed the
LPS-induced decrease in the viability of ATDC5 cells, and also
inhibited the LPS-induced increase in apoptosis, oxidative stress
injury and secretion of proinflammatory cytokines, and activation
of the NF-κB pathways. In addition, the overexpression of NR024118
effectively augmented the LPS-induced changes in the protein
expression levels of Nrf2, HO-1 and NQO1, and activation of the
Nrf2 pathway.
Inflammatory damage of the articular cartilage
serves a vital role in articular cartilage degeneration and OA
progression (15). LPS has been
used to mimic OA in vitro and to examine potential drugs for
OA (16). Therefore, in the
present study, LPS-induced cells were used as a model of OA in
vitro. Consistent with data from a previous study (16), LPS treatment significantly
decreased ATDC5 cell viability, increased cell apoptosis, oxidative
stress and secretion of inflammatory cytokines.
Recently, lncRNAs have been demonstrated to be
involved in a variety of biological processes, and a number of
different types of diseases, including OA. Numerous lncRNAs,
including HOTAIR (17), MALAT1
(18) and MEG3 (19) function to degrade cartilage in
inflammatory injury, and are therefore considered as promising
therapeutic targets of OA. A previous study indicated that shikonin
inhibited the secretion and expression of IL-6 and IL-8 in
rheumatoid arthritis synovial fibroblasts via lncRNA-NR024118
(20). The results of the present
study demonstrated that NR024118 was significantly upregulated in
LPS-treated ATDC5 cells and that the overexpression of NR024118
reversed LPS-induced cell damages in ATDC5 cells.
NF-κB activation exhibits a crucial role in the
LPS-induced inflammatory response (21). LPS stimulation increases the
phosphorylation of IκBβ and NF-κB p65, increasing the transcription
of multiple inflammatory cytokines, including IL-6 and IL-1β
(22). The NF-κB signaling pathway
is generally considered to be a central regulator of the
chondrocyte inflammatory response (23). A previous study has suggested that
lncRNA MALAT1 mitigated the LPS-induced inflammatory damage in
ATDC5 cells by inactivating NF-κB pathways (18). The Nrf2 signaling pathway is an
important endogenous antioxidant pathway. Increasing evidence has
suggested the presence of crosstalk between the NF-κB and Nrf2
signaling pathways. For example, Nrf2 has been indicated to inhibit
NF-κB pathway activation by increasing antioxidant activity and
effectively neutralizing excessive NF-κB activation when exposed to
LPS (24). Nrf2 suppressed lupus
nephritis through neutralizing ROS and by negatively regulating the
NF-κB signaling pathway (25).
Sauchinone inhibits IL-1β-induced catabolism and hypertrophy in
mouse chondrocytes, attenuating OA via the Nrf2/HO-1 and NF-κB
pathways (26). Nrf2 is
hypothesized to suppress the pro-inflammatory pathways mediated by
NF-κB signaling (27,28). The results of the present study
exhibited an activation of the NF-κB and Nrf-2 pathway in ATDC5
cells following treatment with LPS. In the present study, p-IκBβ
and p-p65 expression was increased following LPS stimulation, which
was additionally increased by the overexpression of NR024118. In
addition, NR024118 overexpression effectively augmented the
LPS-induced the protein and mRNA expression levels of Nrf2, HO-1
and NQO1, and activation of the Nrf2 pathway.
In conclusion, the present study demonstrated that
NR024118 levels were downregulated in LPS-induced chondrocytes.
NR024118 overexpression alleviated the LPS-induced cell apoptosis,
oxidative stress and inflammatory injury through the NF-κB and
Nrf-2 signaling pathways. These results suggested that NR024118 may
be a potential target for diagnosis and treatment of patients with
OA.
Acknowledgements
Not applicable.
Funding
The present study was supported by Nanjing Medical
Science and Technique Development Foundation.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XM wrote the paper. XM, JT, and WZ performed the
experiments. XM and JT analyzed the data. YZ designed the
experiments and improved the manuscript. All authors read and
approved the manuscript, and agreed to be accountable for all
aspects of the study in ensuring that the integrity and accuracy of
any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tong W, Geng Y, Huang Y, Shi Y, Xiang S,
Zhang N, Qin L, Shi Q, Chen Q, Dai K and Zhang X: In vivo
identification and induction of articular cartilage stem cells by
inhibiting NF-κB signaling in osteoarthritis. Stem Cells.
33:3125–3137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thomas AC, Hubbard-Turner T, Wikstrom EA
and Palmieri-Smith RM: Epidemiology of posttraumatic
osteoarthritis. J Athl Train. 52:491–496. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mithoefer K: Complex articular cartilage
restoration. Sports Med Arthrosc Rev. 21:31–37. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yucesoy B, Charles LE, Baker B and
Burchfiel CM: Occupational and genetic risk factors for
osteoarthritis: A review. Work. 50:261–273. 2015.PubMed/NCBI
|
|
5
|
Goldring MB: The role of the chondrocyte
in osteoarthritis. Arthritis Rheum. 43:1916–1926. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bay-Jensen AC, Slagboom E, Chen-An P,
Alexandersen P, Qvist P, Christiansen C, Meulenbelt I and Karsdal
MA: Role of hormones in cartilage and joint metabolism:
Understanding an unhealthy metabolic phenotype in osteoarthritis.
Menopause. 20:578–586. 2013.PubMed/NCBI
|
|
7
|
Jiang SD, Lu J, Deng ZH, Li YS and Lei GH:
Long noncoding RNAs in osteoarthritis. Joint Bone Spine.
84:553–556. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li F, Sun J, Huang S, Su G and Pi G:
lncRNA GAS5 overexpression reverses LPS-induced inflammatory injury
and apoptosis through Up-regulating KLF2 expression in ATDC5
chondrocytes. Cell Physiol Biochem. 45:1241–1251. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shih RH, Wang CY and Yang CM: NF-kappaB
signaling pathways in neurological inflammation: A mini review.
Front Mol Neurosci. 8:772015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo C, Urgard E, Vooder T and Metspalu A:
The role of COX-2 and Nrf2/ARE in anti-inflammation and
antioxidative stress: Aging and anti-aging. Med Hypotheses.
77:174–178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahmed SM, Luo L, Namani A, Wang XJ and
Tang X: Nrf2 signaling pathway: Pivotal roles in inflammation.
Biochim Biophys Acta Mol Basis Dis. 1863:585–597. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Q, Niu J, Li H, Ke Y, Li R, Zhang Y
and Lin J: Knee symptomatic osteoarthritis, walking disability,
NSAIDs use and All-cause mortality: Population-based wuchuan
osteoarthritis study. Sci Rep. 7:33092017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saklatvala J: Inflammatory signaling in
cartilage: MAPK and NF-kappaB pathways in chondrocytes and the use
of inhibitors for research into pathogenesis and therapy of
osteoarthritis. Curr Drug Targets. 8:305–313. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang CH, Hsu YM, Chen YC, Lin FH,
Sadhasivam S, Loo ST and Savitha S: Anti-inflammatory effects of
hydrophilic and lipophilic statins with hyaluronic acid against
LPS-induced inflammation in porcine articular chondrocytes. J
Orthop Res. 32:557–565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang C, Wang P, Jiang P, Lv Y, Dong C,
Dai X, Tan L and Wang Z: Upregulation of lncRNA HOTAIR contributes
to IL-1β-induced MMP overexpression and chondrocytes apoptosis in
temporomandibular joint osteoarthritis. Gene. 586:248–253. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pan L, Liu D, Zhao L, Wang L, Xin M and Li
X: Long noncoding RNA MALAT1 alleviates lipopolysaccharide-induced
inflammatory injury by upregulating microRNA-19b in murine
chondrogenic ATDC5 cells. J Cell Biochem. 119:10165–10175. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Z, Chi X, Liu L, Wang Y, Mei X, Yang
Y and Jia T: Long noncoding RNA maternally expressed gene 3
knockdown alleviates lipopolysaccharide-induced inflammatory injury
by up-regulation of miR-203 in ATDC5 cells. Biomed Pharmacother.
100:240–249. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang KY and Chen DL: Shikonin inhibits
inflammatory response in rheumatoid arthritis synovial fibroblasts
via lncRNA-NR024118. Evid Based Complement Alternat Med.
2015:6317372015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang X, Metzger D, Leeman S and Amar S:
LPS-induced TNF-alpha factor (LITAF)-deficient mice express reduced
LPS-induced cytokine: Evidence for LITAF-dependent LPS signaling
pathways. Proc Natl Acad Sci USA. 103:13777–13782. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li L, Liu H, Shi W, Liu H, Yang J, Xu D,
Huang H and Wu L: Insights into the action mechanisms of
traditional Chinese medicine in osteoarthritis. Evid Based
Complement Alternat Med. 2017:51909862017.PubMed/NCBI
|
|
23
|
Ji B, Guo W, Ma H, Xu B, Mu W, Zhang Z,
Amat A and Cao L: Isoliquiritigenin suppresses IL-1β induced
apoptosis and inflammation in chondrocyte-like ATDC5 cells by
inhibiting NF-κB and exerts chondroprotective effects on a mouse
model of anterior cruciate ligament transection. Int J Mol Med.
40:1709–1718. 2017.PubMed/NCBI
|
|
24
|
Chen XY, Dou YX, Luo DD, Zhang ZB, Li CL,
Zeng HF, Su ZR, Xie JH, Lai XP and Li YC: β-Patchoulene from
patchouli oil protects against LPS-induced acute lung injury via
suppressing NF-κB and activating Nrf2 pathways. Int
Immunopharmacol. 50:270–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang T, Tian F, Zheng H, Whitman SA, Lin
Y, Zhang Z, Zhang N and Zhang DD: Nrf2 suppresses lupus nephritis
through inhibition of oxidative injury and the NF-κB-mediated
inflammatory response. Kidney Int. 85:333–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu D, Jin S, Lin Z, Chen R, Pan T, Kang X,
Huang H, Lin C and Pan J: Sauchinone inhibits IL-1β induced
catabolism and hypertrophy in mouse chondrocytes to attenuate
osteoarthritis via Nrf2/HO-1 and NF-κB pathways. Int
Immunopharmacol. 62:181–190. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee DF, Kuo HP, Liu M, Chou CK, Xia W, Du
Y, Shen J, Chen CT, Huo L, Hsu MC, et al: KEAP1 E3 ligase-mediated
downregulation of NF-kappaB signaling by targeting IKKbeta. Mol
Cell. 36:131–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li W, Khor TO, Xu C, Shen G, Jeong WS, Yu
S and Kong AN: Activation of Nrf2-antioxidant signaling attenuates
NFkappaB-inflammatory response and elicits apoptosis. Biochem
Pharmacol. 76:1485–1489. 2008. View Article : Google Scholar : PubMed/NCBI
|