Liver fibrosis presents as a common pathological
process characterized by the overproduction of extracellular matrix
and impaired matrix degradation in the progression of different
liver diseases, such as viral hepatitis and alcoholic liver disease
(1). Hepatic stellate cells
(HSCs) are the predominant non-parenchymal cells which are
activated to repair tissue in response to liver injury; however,
hyperactivation of HSCs results in extracellular matrix deposition
and thus hepatic fibrosis (2). As
an essential component of innate immunity in the liver, natural
killer (NK) cells play a critical role in the removal of pathogens,
including bacteria, virus, and cancer cells (3). Manipulation of NK cell activation
has become a potential cancer immunotherapy, such as adoptive
transfer of allogeneic NK cells, genetic engineered NK cells, NK
cell-targeted chemotherapy and others (4). Several previous reviews have
revealed that NK cells exert antifibrotic functions through their
cytotoxicity against activated HSCs and production of various
cytokines that further intensify this effect (5). Thus, the activation of NK cells is
regarded as a novel therapeutic strategy for hepatic fibrosis
(6). However, dysfunction of NK
cells in the advanced stage of diseases, including liver fibrosis
and tumors, limits its efficacy and clinical application, and the
mechanisms of NK cell dysfunction are still elusive. Further
clarifying the molecular mechanism of NK cells affecting HSCs
during different stages of fibrosis is still required in this
research field. Currently, many treatments have improved fibrosis
at the early stage of this disease, but fail to halt fibrosis
progression in the advanced stage (7). One reason for this is that the
cytotoxic function of NK cells is compromised and HSCs become
resistant to NK cell killing (8).
Therefore, understanding the crosstalk between NK cells and HSCs is
vital for developing novel therapeutic strategies. This review
concisely summarizes the role of HSCs in fibrogenesis, and the
phenotypic and functional characteristics of NK cells in the liver.
It emphasizes the research progression on the crosstalk between NK
cells and HSCs, and the molecular mechanism of NK cell dysfunction
in advanced fibrosis, as well as discusses the therapeutic
potential of the combination of activating NK cell therapies with
TGF-β inhibitors for advanced liver fibrosis.
In order to summarize the interactions between NK
cells and HSCs in liver fibrosis, a PubMed search was performed in
March 2022. Articles containing the following key words were
considered for inclusion: ‘natural killer’ (or ‘NK’) AND ‘hepatic
stellate cells’ (or ‘HSCs’) AND ‘liver fibrosis’ (or ‘hepatic
fibrosis’). Relevant articles were also identified from a manual
search of reference lists within those included. The abstracts of
identified articles were screened and classified for inclusion in
the review. To be included, the article must have described
original data concerning the crosstalk between NK cells and HSCs in
liver fibrosis, and have been published in a peer-reviewed journal
and written in English.
The activation of HSCs is regarded as the initial
event in liver fibrosis. Various pathological agents, such as
viruses, ethanol, and lipids, induce persistent hepatic damage,
which is primarily responsible for HSC activation and
transformation (9). Although
signals that trigger the activation of HSCs are intricate and
remain incompletely elucidated, it has been demonstrated that
several compounds released from dead hepatocytes, including
apoptotic bodies, DNA and inflammasomes can promote HSC activation
(10). Moreover, inflammatory
cells are recruited to lesions and produce numerous cytokines and
growth factors, thereby accelerating HSC proliferation and
differentiation (11). For
instance, the platelet-derived growth factor (PDGF) is the key
factor driving HSC proliferation, while transforming growth factor
(TGF)-β promotes HSC transformation into myofibroblast-like cells
that produce a large amount of collagens, whose deposition beyond
degradation by matrix metalloproteases (MMPs) leads to
extracellular matrix accumulation and thus fibrosis (1).
In healthy livers, HSCs are normally quiescent,
while resident HSCs are activated during liver injury. Activated
HSCs become resistant to Fas ligand (FasL) and tumor necrosis
factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis,
thereby aggravating fibrogenesis (12). Persistent inflammatory and immune
responses in the liver commonly establish an inflammatory
microenvironment that results in the expansion of fibrosis, which
makes it susceptible to progression into cirrhosis and cancer
(13). Therefore, removal of
activated HSCs is essential for a reduction in collagen deposition
and suppression of hepatic fibrosis.
NK cells are cytotoxic large granular lymphocytes
that secrete various cytokines and kill target cells infected by
different pathogens, playing a vital role in the innate immune
system. Human liver NK cells were first described as ‘pit cells’ by
electron microscopy and were found to reside in liver sinusoids
(14,15). Liver-resident NK cells nearly
account for 50% of intrahepatic lymphocytes in the human, and the
percentage of NK cells in the liver is higher than that observed in
the spleen and the peripheral blood (5). Compared with circulating NK cells,
liver NK cells are less mature under the resting condition, but
become more activated in response to pathogenic stimulus and
present higher cytotoxicity (16). These results suggest that the
enrichment of NK cells in the liver might occur in preparation for
exerting particular functions to resist against liver damage. NK
cells are classified into CD56bright and the
CD56dim subsets based on the expression levels of the
activation marker CD69, and CD56bright NK cells display
more responsiveness to cytokines while CD56dim NK cells
harbor more cytotoxicity against pathogens (17). In the liver, the percentage of
these two types of NK cells is equal (18). However, it remains unknown what
regulates the frequency and function of these two types of NK cells
in the liver.
Once hepatocyte damage is triggered by pathogens
such as alcohol and viruses, NK cells are recruited to the hepatic
lesions. Within the hepatic microenvironment, the recruitment and
retention of NK cells are regulated by chemokine receptors
expressed on the surface of NK cells, including CXCR6 and CCR5,
which are engaged in the binding with their cognate ligands
expressed on hepatocytes, thus mediating their migration to the
liver and exerting their immune and cytotoxic functions (19). The main functions of NK cells are
to kill target cells through receptor-ligand interactions and to
secrete a variety of cytokines for assisting the cytotoxicity. In
response to virus infection or tumorigenesis, numerous genomes that
are responsible for encoding activating and inhibitory receptors
are induced to regulate the opposing receptor-ligand signals to
kill target cells (20). When the
activating receptors and inhibitory receptors expressed on NK cells
bind to the corresponding ligands expressed on target cells, NK
cells are either activated or suppressed, respectively (21). Therefore, the activation of NK
cells depends on the balance between the signals from the
stimulatory and inhibitory receptors. Currently, increasing studies
have identified various receptor-ligand signals between NK cells
and their target cells (21), and
several crucial signals will be discussed below. The activated NK
cells kill the target cells through several mechanisms: granule
exocytosis, death receptor-mediated apoptosis (22,23), and cytokine secretion (24). Thus, activation of NK cells is
expected to becoming a promising strategy to halt the progression
of liver diseases.
The interaction between NK cells and HSCs is
manipulated by various stimulatory and inhibitory receptors on NK
cells and the corresponding ligands on HSCs. Among activating NK
cell receptors, NK group 2D (NKG2D) and NKp46 have been extensively
studied in liver fibrosis. It has been reported that expression
levels of NKG2D ligands, including major histocompatibility complex
(MHC) class I polypeptide-related sequence A (MICA) and UL16
binding proteins (ULBPs), are increased in senescent HSCs that are
more susceptible to NK cell killing by granule exocytosis (25). These ligands bind to NKG2D on NK
cells and subsequently induce the activation of NK cells, which
leads to the production of a variety of cytotoxic mediators and
ultimately to HSC death. With the progression of fibrosis, the
expression level of NKp46 is upregulated to kill activated HSCs by
binding to its ligand NCR1 (26).
However, the expression level of NKp46 and cytolytic activity of NK
cells are reduced with the development of fibrosis into the
advanced stage (27,28). This result suggests that
interactions between activating NK cell receptors and ligands are
compromised in the advanced stage of liver fibrosis. In addition,
emerging activating receptors are found to be involved in the NK
cell-mediated killing of HSCs, such as NKp44, killer cell
lectin-like receptor G1 (KLRG1), and Toll-like receptor 9 (TLR-9)
(29–31). Under physiological conditions,
decreased major histocompatibility complex (MHC) I expression
activates HSCs, which are further induced to undergo apoptosis by
NK cells (32). Experimentally,
the genetic deletion of specific inhibitory receptors such as
killer cell immunoglobulin-like receptors (KIRs) and Ly49 increases
the NK toxicity to HSCs and attenuates hepatic fibrosis (33). As a consequence, disruption of the
interaction between inhibitory NK cell receptors and ligands on
HSCs could increase the killing of HSCs and alleviate fibrosis.
HSCs are the predominant non-parenchymal cells in
the liver and regulate the blood flow in hepatic sinusoids. They
are also potent liver-resident antigen presenting cells that
express MHC I molecules to enhance antigen presentation (34,35). According to the missing-self
hypothesis, NK cells target abnormal cells that lack or express low
levels of MHC class I, since MHC I recognizes NK cell inhibitory
receptors and thus dampens their effector function (36). It is generally accepted that the
amount of MHC I expressed on target cells is proportional to the
degree of inhibition. Consequently, NK cells fail to kill quiescent
HSCs under normal condition. In response to liver injury, HSCs are
activated to downregulate the expression of MHC I, which reduces NK
cell suppression and increases the cytotoxicity of NK cells against
activated HSCs (32,33). Compared with activated HSCs,
quiescent HSCs generate less retinoic acid and have decreased
expression of retinoic acid early transcript-1 (RAET1E), a ligand
for activating NK cell receptors, thereby preventing them from NK
cell-mediated killing (37).
Generally, the activation of HSCs induced by
hepatocyte damage is attributed to increased NK cell stimulation
and decreased NK cell inhibition (3). Activated NK cells produce a large
amount of perforin and granzyme and induce activated HSC apoptosis
(32). At the early stage of
fibrosis, NK cells are recruited to the liver and harbor potent
degranulation activity that are responsible for the removal of
activated HSCs, thus delaying the progression of fibrogenesis
(38). With the development of
this disease, the degranulation activity of NK cells is impaired in
the advanced fibrosis stage (39). These findings suggest that
activated HSCs are sensitive to NK killing at the initial stage but
become blunt in the advanced stage of fibrosis. In addition,
activation of HSCs upregulates the expression of TRAIL receptor and
Fas, which interact with NK cells by TRAIL/TRAIL receptor and
Fas/FasL signaling, leading to enhanced cytotoxicity of NK cells
against activated HSCs. For example, activated HSCs highly express
RAE-1, and are killed by activated NK cells through TRAIL-mediated
apoptosis (40). Increased TRAIL
expression on NK cells promotes the killing attack against
activated HSCs (41,42). A clinical investigation also
confirmed that NK cells from HCV-infected patients induced
apoptosis of activated HSCs in the TRAIL and FasL-dependent manners
(43). These results are
consistent with the fact that administration of TRAIL induces
apoptosis of activated HSCs and thus alleviates liver fibrosis
(44). Furthermore, activated NK
cells secrete various antifibrotic cytokines to induce activated
HSC death. Several studies have revealed that IFN-γ released from
NK cells mediates cell cycle arrest and proliferation inhibition of
activated HSCs (41,45,46).
Senescence is a cellular process that irreversibly
blocks cell proliferation. Senescent HSCs have reduced viability,
decreased production of extracellular matrix component, enhanced
secretion of extracellular matrix-degrading enzymes, and increased
immune surveillance, resulting in limited fibrosis (47). The senescence of activated HSCs
renders them susceptible to killing by NK cells, thereby
facilitating the resolution of fibrosis (48). In liver injury, intrahepatic NK
cells are activated to produce high levels of interleukin (IL)-10
and IL-22, which are able to trigger the innate immune response,
protection from damage, and regeneration (49,50). Several studies have shown that
both IL-10 and IL-22 promote the senescence of activated HSCs via
the STAT3/p53/p21 pathway, attenuating liver fibrosis (51–53). A recent study demonstrated that
activation of this pathway accelerates HSC senescence and makes
them more vulnerable to being eliminated by NK cells (54). These findings indicate that NK
cells produce cytokines to facilitate the senescence of activated
HSCs and further to promote cell death. In addition, senescent HSCs
induced by the natural product curcumin through the PPARγ/P53
signaling, can be easily removed by NK cells (25,55), suggesting that curcumin serves as
a senescence inducer to play a protective role in fibrosis. The
mechanism by which NK cells kill senescent HSCs is still unclear,
but a study revealed that the lack of granule exocytosis in NK
cells resulted in excessive accumulation of senescent HSCs and
aggravated liver fibrosis, indicating that NK cells kill senescent
HSCs by granule exocytosis; moreover, senescent HSCs with high Dcr2
expression are killed by NK cells through granule exocytosis
(56). Therefore, further
clarifying the mechanisms of the killing of senescent HSCs by NK
cells may provide an effective therapeutic strategy for liver
fibrosis.
The initial activation of NK cells in the early
stage of fibrosis is believed to eliminate activated HSCs and
restrict fibrogenesis; however, these effects are weakened and HSCs
become resistant to the killing of NK cells in the advanced stage.
Previous research has revealed that NK cells from patients with
cirrhosis directly contact cultured HSCs and are subsequently
engulfed by HSCs via the Rac1 and Cdc42 pathways (57). The phagocytosis of NK cells by
HSCs is referred as emperipolesis that is mediated by TGF-β, an
immunosuppressive cytokine derived from HSCs. This process promotes
NK cell apoptosis and contributes to NK cell depletion, impairing
the anti-fibrosis capacity of NK cells (58). Phagocytosis of apoptotic NK cells
further makes HSCs resistant to FasL and TRAIL-induced apoptosis
via the JAK/STAT and Akt/NF-kB-dependent pathways (12). These findings suggest that
TGF-β-mediated emperipolesis of NK cells exacerbates liver
fibrosis. Indeed, TGF-β is also known to suppress NK cell-induced
cytotoxicity and cytokine production (59). Data from a pre-clinical setting
demonstrated that overproduction of TGF-β by HSCs renders them
resistant to NK cell killing and inhibits the anti-fibrotic effect
of NK cells in advanced liver fibrosis where NK cells have reduced
expression of NKG2D, TRAIL and interferon (IFN)-γ (8,60).
The mechanisms by which TGF-β impairs the cytotoxicity of NK cells
against HSCs remain to be investigated. A recent study has shown
that the defective capacity to produce IFN-γ in NK cells is
reversed by TGF-β blockade in patients with chronic hepatitis B
virus infection (61). It can be
assumed that the increased TGF-β levels inhibit the production of
IFN-γ in NK cells and makes HSCs resistant to IFN-γ-induced cell
cycle arrest and apoptosis in the liver with advanced fibrosis.
Moreover, a series of studies have uncovered that elevated TGF-β
suppresses the antifibrotic function of NK cells via downregulation
of NKG2D and 2B4 surface expression (62–64), thereby reducing the activation of
NK cells. Therefore, TGF-β derived from HSCs mediates the
resistance to NK cell cytotoxicity in advanced liver fibrosis.
Blockade of TGF-β might provide a promising therapeutic strategy
for this disease.
With liver disease progression to fibrosis, the
effector function of NK cells is activated to eliminate HSCs at the
early stage but exhausted at the advanced stage. Therefore, the
initial intensification of the antifibrotic function of NK cells
can halt the development of liver fibrosis. Experimentally, several
studies have unveiled that upregulation of the expression level of
NKG2D in NK cells by natural products, such as Mycelium
cordyceps sinensis and Salvia miltiorrhiza, alleviates
liver fibrosis (65,66). Using siRNA to downregulate the
expression of KIR also limits fibrosis (33). These findings imply that
modulation of the expression of activating and inhibitory NK cell
receptors may be a promising strategy to activate NK cells. In
addition, an enhancement of NK cell-derived antifibrotic cytokines
has been confirmed to be effective in killing HSCs. For instance,
the herbal agent Yu Gan Long was found to increase the expression
level of IFN-γ and facilitate extracellular matrix degradation
(67). Exogenous IFN-γ
supplementation delivered to HSCs directly elevates the
antifibrotic potency of NK cells (68). Thus, activation of NK cells is
recognized as a novel therapeutic strategy for treatment of liver
fibrosis (5). However, in the
advanced fibrosis stage, the expression of activating NK cell
receptors, the production of IFN-γ, the degranulation activity, and
the cytotoxic function of NK cells are diminished after interaction
with activated HSCs (39,69,70), suggesting a functional exhaustion
of NK cells. Unfortunately, as concluded above, HSCs become
resistant to these exhausted NK cells in the advanced stage of
disease. Only the activation of NK cells is insufficient to recover
their cytotoxic function in advance fibrosis. A recent study found
that fasudil facilitates the apoptosis and inhibits the
proliferation of HSCs by decreasing TGF-β (71). Thus, NK cell activation combined
with TGF-β blockade may be effective to restrict fibrogenesis in
the advanced stage of the disease.
As the initial event in fibrogenesis, HSCs are
activated to tissue repair in liver injury, but hyperactivation of
HSCs produces excessive collagens and contributes to fibrosis.
Hepatic NK cells, an essential component of the innate immunity,
are activated to eliminate HSCs through receptor-ligand
interactions and to produce various antifibrotic cytokines to
assist their cytotoxicity. These effects are modulated by both
activating and inhibitory NK cell receptors. Under physiological
condition, quiescent HSCs fail to be targeted by NK cells, but
activated and senescent HSCs can be killed by activated NK cells in
the early stage of fibrosis through granule exocytosis, death
receptor-mediated pathways, and production of antifibrotic
cytokines; however, in the advanced stage, the cytotoxicity of NK
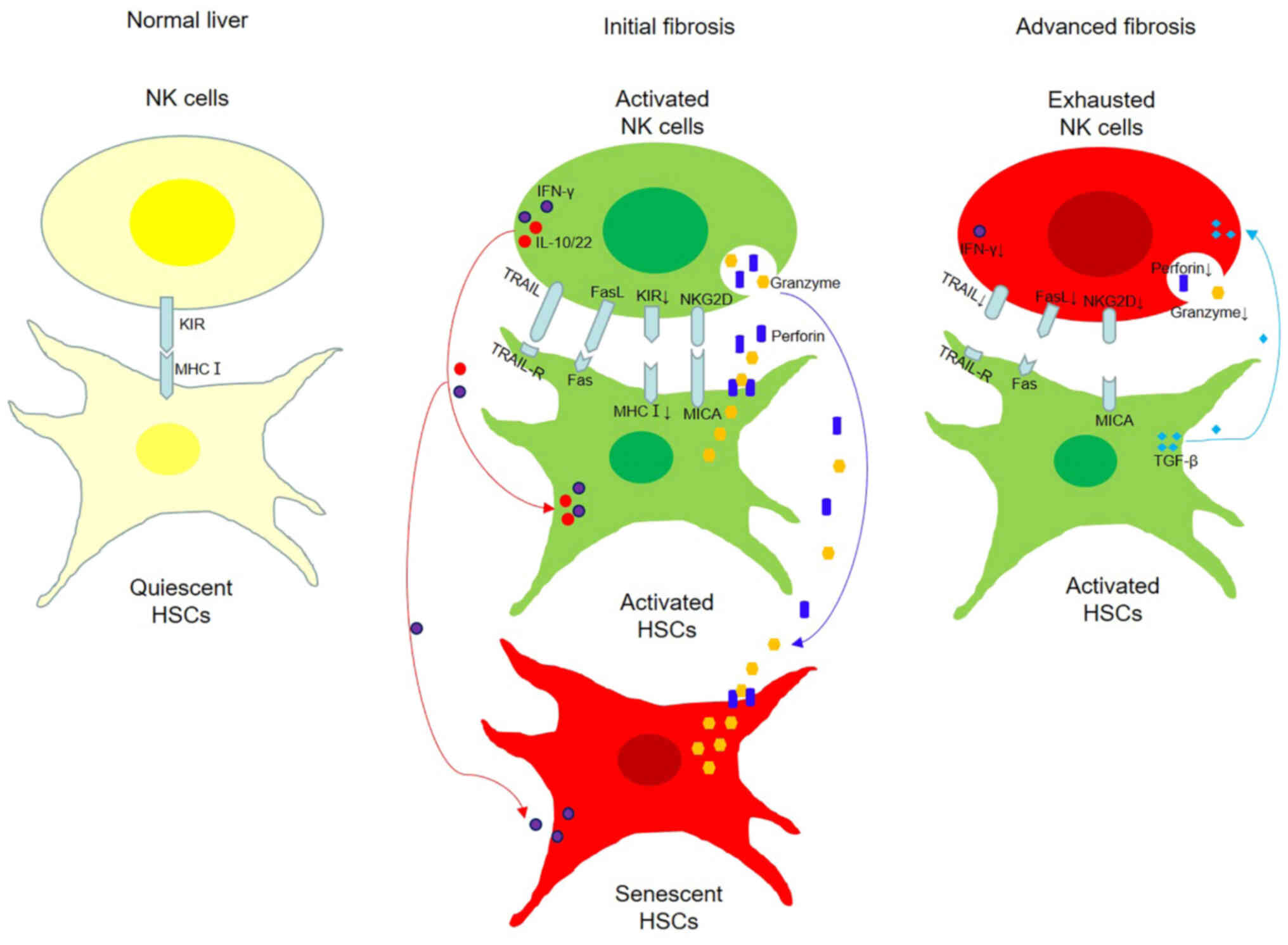
cells against HSCs is defective (Fig.
1). A summary of the crosstalk between HSCs and NK cells in
fibrosis of various liver diseases is described in Table I (25,32,33,37,40,43,46,54,56–58,60).
Experimentally, a series of studies demonstrated
that activating NK cells in the initial stage of disease postpone
the development of liver fibrosis, but fail to halt fibrogenesis in
the advanced stage (8,38,39,60). The mechanism of dysfunction in NK
cells is still elusive, but it is partially attributed to the fact
that HSCs become resistant to exhausted NK cells in a
TGF-β-dependent manner. This may provide an additional clue to
explain why activating NK cells fail in progressive fibrosis. In
this content, activating NK cell therapies combined with a TGF-β
inhibitor will likely represent a promising therapeutic strategy in
the disease, and merits further verification in clinical
investigations. In addition, the cytotoxic function of NK cells is
regulated by a complex network of activating and inhibitory
receptors. Therefore, further clarification of NK cell receptors
and their cytotoxic functions will provide new insight in the
etiology of fibrogenesis, as potential therapies for liver
fibrosis.
Not applicable.
This work was funded by the National Natural Science Foundation
of China (grant no. 81973601) and TCM Research Projects of
Heilongjiang Province (grant nos. ZHY18-029, ZHY19-027, ZHY19-062
and ZHY2020-041).
All information provided in this review is
documented by relevant references.
XY and BW designed the review. YZ, YW and WS wrote
the review in light of the literature findings. All authors read
and approved the final manuscript for publication. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare no competing interests.
|
1
|
Gan J, Mao XR, Zheng SJ and Li JF:
Invariant natural killer T cells: Not to be ignored in liver
disease. J Dig Dis. 22:136–142. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yan Y, Zeng J, Xing L and Li C: Extra- and
intra-cellular mechanisms of hepatic stellate cell activation.
Biomedicines. 9:10142021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Highton AJ, Schuster IS, Degli-Esposti MA
and Altfeld M: The role of natural killer cells in liver
inflammation. Semin Immunopathol. 43:519–533. 2021. View Article : Google Scholar
|
|
4
|
St-Pierre F, Bhatia S and Chandra S:
Harnessing natural killer cells in cancer immunotherapy: A review
of mechanisms and novel therapies. Cancers (Basel). 13:19882021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zheng M, Sun H and Tian Z: Natural killer
cells in liver diseases. Front Med. 12:269–279. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gao B, Radaeva S and Jeong WI: Activation
of natural killer cells inhibits liver fibrosis: A novel strategy
to treat liver fibrosis. Expert Rev Gastroenterol Hepatol.
1:173–180. 2007. View Article : Google Scholar
|
|
7
|
Schuppan D, Ashfaq-Khan M, Yang AT and Kim
YO: Liver fibrosis: Direct antifibrotic agents and targeted
therapies. Matrix Biol. 68-69:435–451. 2018. View Article : Google Scholar
|
|
8
|
Jeong WI, Park O, Suh YG, Byun JS, Park
SY, Choi E, Kim JK, Ko H, Wang H, Miller AM and Gao B: Suppression
of innate immunity (natural killer cell/interferon-gamma) in the
advanced stages of liver fibrosis in mice. Hepatology.
53:1342–1351. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Roehlen N, Crouchet E and Baumert TF:
Liver fibrosis: Mechanistic concepts and therapeutic perspectives.
Cells. 9:8752020. View Article : Google Scholar
|
|
10
|
Wree A, McGeough MD, Inzaugarat ME, Eguchi
A, Schuster S, Johnson CD, Peña CA, Geisler LJ, Papouchado BG,
Hoffman HM and Feldstein AE: NLRP3 inflammasome driven liver injury
and fibrosis: Roles of IL-17 and TNF in mice. Hepatology.
67:736–749. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wan M, Han J, Ding L, Hu F and Gao P:
Novel immune subsets and related cytokines: Emerging players in the
progression of liver fibrosis. Front Med. 8:6048942021. View Article : Google Scholar
|
|
12
|
Jiang JX, Mikami K, Venugopal S, Li Y and
Török NJ: Apoptotic body engulfment by hepatic stellate cells
promotes their survival by the JAK/STAT and Akt/NF-kappaB-dependent
pathways. J Hepatol. 51:139–148. 2009. View Article : Google Scholar
|
|
13
|
Tanwar S, Rhodes F, Srivastava A,
Trembling PM and Rosenberg WM: Inflammation and fibrosis in chronic
liver diseases including non-alcoholic fatty liver disease and
hepatitis C. World J Gastroenterol. 26:109–133. 2020. View Article : Google Scholar
|
|
14
|
Luo DZ, Vermijlen D, Ahishali B, Triantis
V, Plakoutsi G, Braet F, Vanderkerken K and Wisse E: On the cell
biology of pit cells, the liver-specific NK cells. World J
Gastroenterol. 6:1–11. 2000. View Article : Google Scholar
|
|
15
|
Wisse E, van't Noordende JM, van der
Meulen J and Daems WT: The pit cell: Description of a new type of
cell occurring in rat liver sinusoids and peripheral blood. Cell
Tissue Res. 173:423–435. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang L, Peng H, Zhou J, Chen Y, Wei H, Sun
R, Yokoyama WM and Tian Z: Differential phenotypic and functional
properties of liver-resident NK cells and mucosal ILC1s. J
Autoimmun. 67:29–35. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y and Zhang C: The Roles of
Liver-resident lymphocytes in liver diseases. Front Immunol.
10:15822019. View Article : Google Scholar
|
|
18
|
Lunemann S, Martrus G, Goebels H, Kautz T,
Langeneckert A, Salzberger W, Koch M, J Bunders M, Nashan B, van
Gisbergen KPJM and Altfeld M: Hobit expression by a subset of human
liver-resident CD56bright Natural Killer cells. Sci Rep.
7:66762017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hudspeth K, Donadon M, Cimino M, Pontarini
E, Tentorio P, Preti M, Hong M, Bertoletti A, Bicciato S,
Invernizzi P, et al: Human liver-resident CD56(bright)/CD16(neg) NK
cells are retained within hepatic sinusoids via the engagement of
CCR5 and CXCR6 pathways. J Autoimmun. 66:40–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang C, Hu Y, Xiao W and Tian Z: Chimeric
antigen receptor- and natural killer cell receptor-engineered
innate killer cells in cancer immunotherapy. Cell Mol Immunol.
18:2083–2100. 2021. View Article : Google Scholar
|
|
21
|
Obajdin J, Davies DM and Maher J:
Engineering of chimeric natural killer cell receptors to develop
precision adoptive immunotherapies for cancer. Clin Exp Immunol.
202:11–27. 2020. View Article : Google Scholar
|
|
22
|
Vrazo AC, Hontz AE, Figueira SK, Butler
BL, Ferrell JM, Binkowski BF, Li J and Risma KA: Live cell
evaluation of granzyme delivery and death receptor signaling in
tumor cells targeted by human natural killer cells. Blood.
126:e1–e10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saparbay J, Tanaka Y, Tanimine N, Ohira M
and Ohdan H: Everolimus enhances TRAIL-mediated anti-tumor activity
of liver resident natural killer cells in mice. Transpl Int.
33:229–243. 2020. View Article : Google Scholar
|
|
24
|
Choi WM, Ryu T, Lee JH, Shim YR, Kim MH,
Kim HH, Kim YE, Yang K, Kim K, Choi SE, et al: Metabotropic
glutamate receptor 5 in natural killer cells attenuates liver
fibrosis by exerting cytotoxicity to activated stellate cells.
Hepatology. 74:2170–2185. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jin H, Jia Y, Yao Z, Huang J, Hao M, Yao
S, Lian N, Zhang F, Zhang C, Chen X, et al: Hepatic stellate cell
interferes with NK cell regulation of fibrogenesis via curcumin
induced senescence of hepatic stellate cell. Cell Signal. 33:79–85.
2017. View Article : Google Scholar
|
|
26
|
Gur C, Doron S, Kfir-Erenfeld S, Horwitz
E, Abu-Tair L, Safadi R and Mandelboim O: NKp46-mediated killing of
human and mouse hepatic stellate cells attenuates liver fibrosis.
Gut. 61:885–893. 2012. View Article : Google Scholar
|
|
27
|
Li W, Jiang Y, Wang X, Jin J, Qi Y, Chi X,
Zhang H, Feng X and Niu J: Natural killer p46 controls hepatitis B
virus replication and modulates liver inflammation. PLoS One.
10:e01358742015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nattermann J, Feldmann G, Ahlenstiel G,
Langhans B, Sauerbruch T and Spengler U: Surface expression and
cytolytic function of natural killer cell receptors is altered in
chronic hepatitis C. Gut. 55:869–877. 2006. View Article : Google Scholar
|
|
29
|
Nel I, Lucar O, Petitdemange C, Béziat V,
Lapalus M, Bédossa P, Debré P, Asselah T, Marcellin P and Vieillard
V: Accumulation of intrahepatic TNF-α-producing NKp44+ NK cells
correlates with liver fibrosis and viral load in chronic HCV
infection. Medicine (Baltimore). 95:e36782016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wijaya RS, Read SA, Schibeci S, Eslam M,
Azardaryany MK, El-Khobar K, van der Poorten D, Lin R, Yuen L, Lam
V, et al: KLRG1+ natural killer cells exert a novel antifibrotic
function in chronic hepatitis B. J Hepatol. 71:252–264. 2019.
View Article : Google Scholar
|
|
31
|
Abu-Tair L, Axelrod JH, Doron S, Ovadya Y,
Krizhanovsky V, Galun E, Amer J and Safadi R: Natural killer
cell-dependent anti-fibrotic pathway in liver injury via Toll-like
receptor-9. PLoS One. 8:e825712013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cai X, Wang J, Wang J, Zhou Q, Yang B, He
Q and Weng Q: Intercellular crosstalk of hepatic stellate cells in
liver fibrosis: New insights into therapy. Pharmacol Res.
155:1047202020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Muhanna N, Abu Tair L, Doron S, Amer J,
Azzeh M, Mahamid M, Friedman S and Safadi R: Amelioration of
hepatic fibrosis by NK cell activation. Gut. 60:90–98. 2011.
View Article : Google Scholar
|
|
34
|
Schölzel K, Schildberg FA, Welz M, Börner
C, Geiger S, Kurts C, Heikenwälder M, Knolle PA and Wohlleber D:
Transfer of MHC-class-I molecules among liver sinusoidal cells
facilitates hepatic immune surveillance. J Hepatol. 61:600–608.
2014. View Article : Google Scholar
|
|
35
|
Arriola Benitez PC, Pesce Viglietti AI,
Elizalde MM, Giambartolomei GH, Quarleri JF and Delpino MV: Hepatic
stellate cells and hepatocytes as liver Antigen-presenting cells
during B. abortus Infection. Pathogens. 9:5272020. View Article : Google Scholar
|
|
36
|
Chen Y, Lu D, Churov A and Fu R: Research
progress on NK cell receptors and their signaling pathways.
Mediators Inflamm. 2020:64370572020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Radaeva S, Wang L, Radaev S, Jeong WI,
Park O and Gao B: Retinoic acid signaling sensitizes hepatic
stellate cells to NK cell killing via upregulation of NK cell
activating ligand RAE1. Am J Physiol Gastrointest Liver Physiol.
293:G809–G816. 2007. View Article : Google Scholar
|
|
38
|
Park O, Jeong WI, Wang L, Wang H, Lian ZX,
Gershwin ME and Gao B: Diverse roles of invariant natural killer T
cells in liver injury and fibrosis induced by carbon tetrachloride.
Hepatology. 49:1683–1694. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fugier E, Marche H, Thelu MA, Macek
Jílková Z, Van Campenhout N, Dufeu-Duchesne T, Leroy V, Zarski JP,
Sturm N, Marche PN and Jouvin-Marche E: Functions of liver natural
killer cells are dependent on the severity of liver inflammation
and fibrosis in chronic hepatitis C. PLoS One. 9:e956142014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Radaeva S, Sun R, Jaruga B, Nguyen VT,
Tian Z and Gao B: Natural killer cells ameliorate liver fibrosis by
killing activated stellate cells in NKG2D-dependent and tumor
necrosis factor-related apoptosis-inducing ligand-dependent
manners. Gastroenterology. 130:435–452. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jeong WI, Park O, Radaeva S and Gao B:
STAT1 inhibits liver fibrosis in mice by inhibiting stellate cell
proliferation and stimulating NK cell cytotoxicity. Hepatology.
44:1441–1451. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ma PF, Gao CC, Yi J, Zhao JL, Liang SQ,
Zhao Y, Ye YC, Bai J, Zheng QJ, Dou KF, et al: Cytotherapy with
M1-polarized macrophages ameliorates liver fibrosis by modulating
immune microenvironment in mice. J Hepatol. 67:770–779. 2017.
View Article : Google Scholar
|
|
43
|
Glassner A, Eisenhardt M, Kramer B, Körner
C, Coenen M, Sauerbruch T, Spengler U and Nattermann J: NK cells
from HCV-infected patients effectively induce apoptosis of
activated primary human hepatic stellate cells in a TRAIL-, FasL-
and NKG2D-dependent manner. Lab Invest. 92:967–977. 2012.
View Article : Google Scholar
|
|
44
|
Oh Y, Park O, Swierczewska M, Hamilton JP,
Park JS, Kim TH, Lim SM, Eom H, Jo DG, Lee CE, et al: Systemic
PEGylated TRAIL treatment ameliorates liver cirrhosis in rats by
eliminating activated hepatic stellate cells. Hepatology.
64:209–223. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rockey DC, Maher JJ, Jarnagin WR, Gabbiani
G and Friedman SL: Inhibition of rat hepatic lipocyte activation in
culture by interferon-gamma. Hepatology. 16:776–784. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hou X, Yu F, Man S, Huang D, Zhang Y, Liu
M, Ren C and Shen J: Negative regulation of Schistosoma japonicum
egg-induced liver fibrosis by natural killer cells. PLoS Negl Trop
Dis. 6:e14562012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Parola M and Pinzani M: Liver fibrosis:
Pathophysiology, pathogenetic targets and clinical issues. Mol
Aspects Med. 65:37–55. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang M, Serna-Salas S, Damba T, Borghesan
M, Demaria M and Moshage H: Hepatic stellate cell senescence in
liver fibrosis: Characteristics, mechanisms and perspectives. Mech
Ageing Dev. 199:1115722021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fan Y, Zhang W, Wei H, Sun R, Tian Z and
Chen Y: Hepatic NK cells attenuate fibrosis progression of
non-alcoholic steatohepatitis in dependent of CXCL10-mediated
recruitment. Liver Int. 40:598–608. 2020. View Article : Google Scholar
|
|
50
|
Wu Y, Min J, Ge C, Shu J, Tian D, Yuan Y
and Zhou D: Interleukin 22 in liver injury, inflammation and
cancer. Int J Biol Sci. 16:2405–2413. 2020. View Article : Google Scholar
|
|
51
|
Huang YH, Chen MH, Guo QL, Chen YX, Zhang
LJ, Chen ZX and Wang XZ: Interleukin-10 promotes primary rat
hepatic stellate cell senescence by upregulating the expression
levels of p53 and p21. Mol Med Rep. 17:5700–5707. 2018.PubMed/NCBI
|
|
52
|
Huang YH, Chen MH, Guo QL, Chen ZX, Chen
QD and Wang XZ: Interleukin-10 induces senescence of activated
hepatic stellate cells via STAT3-p53 pathway to attenuate liver
fibrosis. Cell Signal. 66:1094452020. View Article : Google Scholar
|
|
53
|
Kong X, Feng D, Wang H, Hong F, Bertola A,
Wang FS and Gao B: Interleukin-22 induces hepatic stellate cell
senescence and restricts liver fibrosis in mice. Hepatology.
56:1150–1159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen J, Xu T, Zhu D, Wang J, Huang C, Lyu
L, Hu B, Sun W and Duan Y: Egg antigen p40 of Schistosoma japonicum
promotes senescence in activated hepatic stellate cells by
activation of the STAT3/p53/p21 pathway. Cell Death Dis.
7:e23152016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Jin H, Lian N, Zhang F, Chen L, Chen Q, Lu
C, Bian M, Shao J, Wu L and Zheng S: Activation of PPARγ/P53
signaling is required for curcumin to induce hepatic stellate cell
senescence. Cell Death Dis. 7:e21892016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sagiv A, Biran A, Yon M, Simon J, Lowe SW
and Krizhanovsky V: Granule exocytosis mediates immune surveillance
of senescent cells. Oncogene. 32:1971–1977. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Muhanna N, Doron S, Wald O, Horani A, Eid
A, Pappo O, Friedman SL and Safadi R: Activation of hepatic
stellate cells after phagocytosis of lymphocytes: A novel pathway
of fibrogenesis. Hepatology. 48:963–977. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shi J, Zhao J, Zhang X, Cheng Y, Hu J, Li
Y, Zhao X, Shang Q, Sun Y, Tu B, et al: Activated hepatic stellate
cells impair NK cell anti-fibrosis capacity through a
TGF-β-dependent emperipolesis in HBV cirrhotic patients. Sci Rep.
7:445442017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yu J, Wei M, Becknell B, Trotta R, Liu S,
Boyd Z, Jaung MS, Blaser BW, Sun J, Benson DM Jr, et al: Pro- and
antiinflammatory cytokine signaling: Reciprocal antagonism
regulates interferon-gamma production by human natural killer
cells. Immunity. 24:575–590. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jeong WI, Park O and Gao B: Abrogation of
the antifibrotic effects of natural killer cells/interferon-gamma
contributes to alcohol acceleration of liver fibrosis.
Gastroenterology. 134:248–258. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Peppa D, Micco L, Javaid A, Kennedy PT,
Schurich A, Dunn C, Pallant C, Ellis G, Khanna P, Dusheiko G, et
al: Blockade of immunosuppressive cytokines restores NK cell
antiviral function in chronic hepatitis B virus infection. PLoS
Pathog. 6:e10012272010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Dasgupta S, Bhattacharya-Chatterjee M,
O'Malley BW Jr and Chatterjee SK: Inhibition of NK cell activity
through TGF-beta 1 by down-regulation of NKG2D in a murine model of
head and neck cancer. J Immunol. 175:5541–5550. 2005. View Article : Google Scholar
|
|
63
|
Sene D, Levasseur F, Abel M, Lambert M,
Camous X, Hernandez C, Pène V, Rosenberg AR, Jouvin-Marche E,
Marche PN, et al: Hepatitis C virus (HCV) evades NKG2D-dependent NK
cell responses through NS5A-mediated imbalance of inflammatory
cytokines. PLoS Pathog. 6:e10011842010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Sun C, Fu B, Gao Y, Liao X, Sun R, Tian Z
and Wei H: TGF-β1 down-regulation of NKG2D/DAP10 and 2B4/SAP
expression on human NK cells contributes to HBV persistence. PLoS
Pathog. 8:e10025942012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Peng Y, Huang K, Shen L, Tao YY and Liu
CH: Cultured mycelium cordyceps sinensis alleviates CCl4-induced
liver inflammation and fibrosis in mice by activating hepatic
natural killer cells. Acta Pharmacol Sin. 37:204–216. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Peng Y, Yang T, Huang K, Shen L, Tao Y and
Liu C: Salvia miltiorrhiza ameliorates liver fibrosis by activating
hepatic natural killer cells in vivo and in vitro. Front Pharmacol.
9:7622018. View Article : Google Scholar
|
|
67
|
Xia Y, Yu B, Ma C, Tu Y, Zhai L, Yang Y,
Liu D, Liu Y, Wu H, Dan H and You P: Yu Gan Long reduces rat liver
fibrosis by blocking TGF-β 1/Smad pathway and modulating the
immunity. Biomed Pharmacother. 106:1332–1338. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
van Dijk F, Olinga P, Poelstra K and
Beljaars L: Targeted therapies in liver fibrosis: Combining the
best parts of Platelet-Derived growth factor BB and interferon
gamma. Front Med (Lausanne). 2:722015.PubMed/NCBI
|
|
69
|
Polo ML, Ghiglione YA, Salido JP, Urioste
A, Poblete G, Sisto AE, Martinez A, Rolón MJ, Ojeda DS, Cahn PE, et
al: Liver cirrhosis in HIV/HCV-coinfected individuals is related to
NK cell dysfunction and exhaustion, but not to an impaired NK cell
modulation by CD4+ T-cells. J Int AIDS Soc.
22:e253752019. View Article : Google Scholar
|
|
70
|
Li Y, Wang JJ, Gao S, Liu Q, Bai J, Zhao
XQ, Hao YH, Ding HH, Zhu F, Yang DL and Zhao XP: Decreased
peripheral natural killer cells activity in the immune activated
stage of chronic hepatitis B. PLoS One. 9:e869272014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Han QJ, Mu YL, Zhao HJ, Zhao RR, Guo QJ,
Su YH and Zhang J: Fasudil prevents liver fibrosis via activating
natural killer cells and suppressing hepatic stellate cells. World
J Gastroenterol. 27:3581–3594. 2021. View Article : Google Scholar
|